

Molecular bases for sensitivity to acetyl-coenzyme A carboxylase inhibitors in black-grass
- PMID: 15579665
- PMCID: PMC1065379
- DOI: 10.1104/pp.104.046144
Molecular bases for sensitivity to acetyl-coenzyme A carboxylase inhibitors in black-grass
Abstract
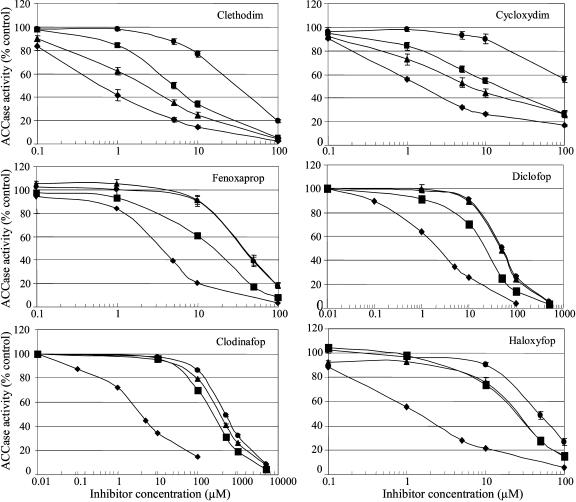
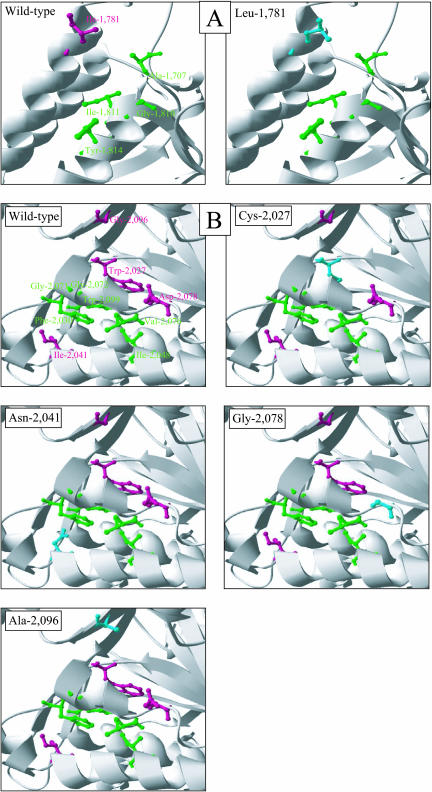
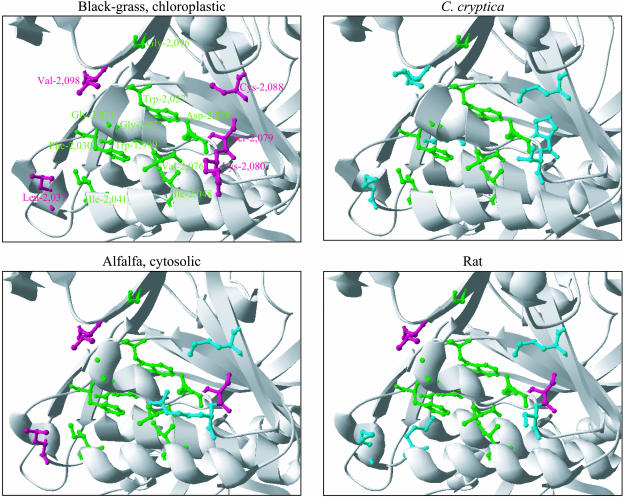
In grasses, residues homologous to residues Ile-1,781 and Ile-2,041 in the carboxyl-transferase (CT) domain of the chloroplastic acetyl-coenzyme A (CoA) carboxylase (ACCase) from the grass weed black-grass (Alopecurus myosuroides [Huds.]) are critical determinants for sensitivity to two classes of ACCase inhibitors, aryloxyphenoxypropionates (APPs) and cyclohexanediones. Using natural mutants of black-grass, we demonstrated through a molecular, biological, and biochemical approach that residues Trp-2,027, Asp-2,078, and Gly-2,096 are also involved in sensitivity to ACCase inhibitors. In addition, residues Trp-2,027 and Asp-2,078 are very likely involved in CT activity. Using three-dimensional modeling, we found that the side chains of the five residues are adjacent, located at the surface of the inside of the cavity of the CT active site, in the vicinity of the binding site for APPs. Residues 1,781 and 2,078 are involved in sensitivity to both APPs and cyclohexanediones, whereas residues 2,027, 2,041, and 2,096 are involved in sensitivity to APPs only. This suggests that the binding sites for these two classes of compounds are overlapping, although distinct. Comparison of three-dimensional models for black-grass wild-type and mutant CTs and for CTs from organisms with contrasted sensitivity to ACCase inhibitors suggested that inhibitors fitting into the cavity of the CT active site of the chloroplastic ACCase from grasses to reach their active sites may be tight. The three-dimensional shape of this cavity is thus likely of high importance for the efficacy of ACCase inhibitors.
Figures
References
-
- Burton JD, Gronwald JW, Keith RA, Somers DA, Gegenbach BG, Wyse DL (1991) Kinetics of inhibition of acetyl-coenzyme A carboxylase by sethoxydim and haloxyfop. Pestic Biochem Physiol 39: 100–109 - PubMed
-
- Christoffers MJ, Berg ML, Messersmith CG (2000) Analysis of acetyl-CoA carboxylase gene sequences from fenoxaprop-P-resistant wild oat biotypes (abstract no. 121). Proc North Cent Weed Sci Soc 55: 67
-
- Christoffers MJ, Berg ML, Messersmith CG (2002) An isoleucine to leucine mutation in acetyl-CoA carboxylase confers herbicide resistance in wild oat. Genome 45: 1049–1056 - PubMed
-
- Christopher JT, Holtum JAM (1998) The dicotyledonous species Erodium moschatum (L) L'Hér. ex. Aiton is sensitive to haloxyfop herbicide due to herbicide-sensitive acetyl-coenzyme A carboxylase. Planta 207: 275–279
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
