

Vacuolar ATPase regulates zymogen activation in pancreatic acini
- PMID: 15582989
- PMCID: PMC2846595
- DOI: 10.1074/jbc.M413513200
Vacuolar ATPase regulates zymogen activation in pancreatic acini
Abstract
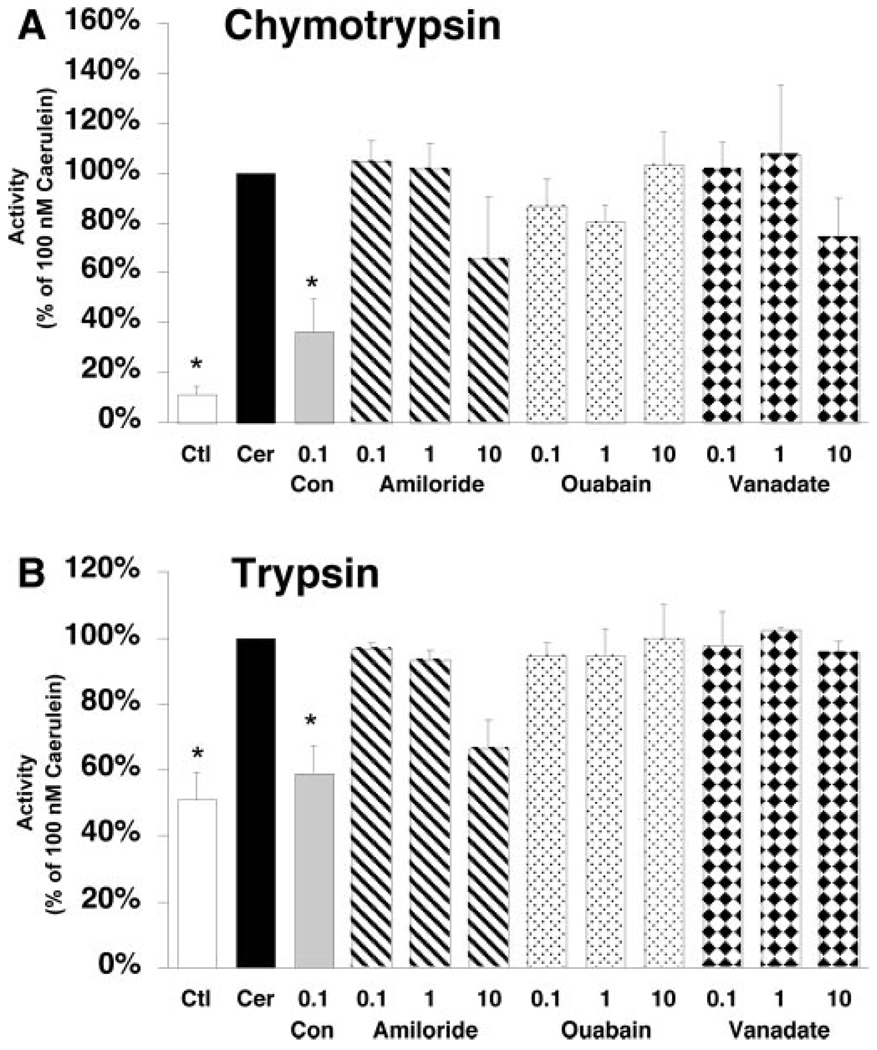
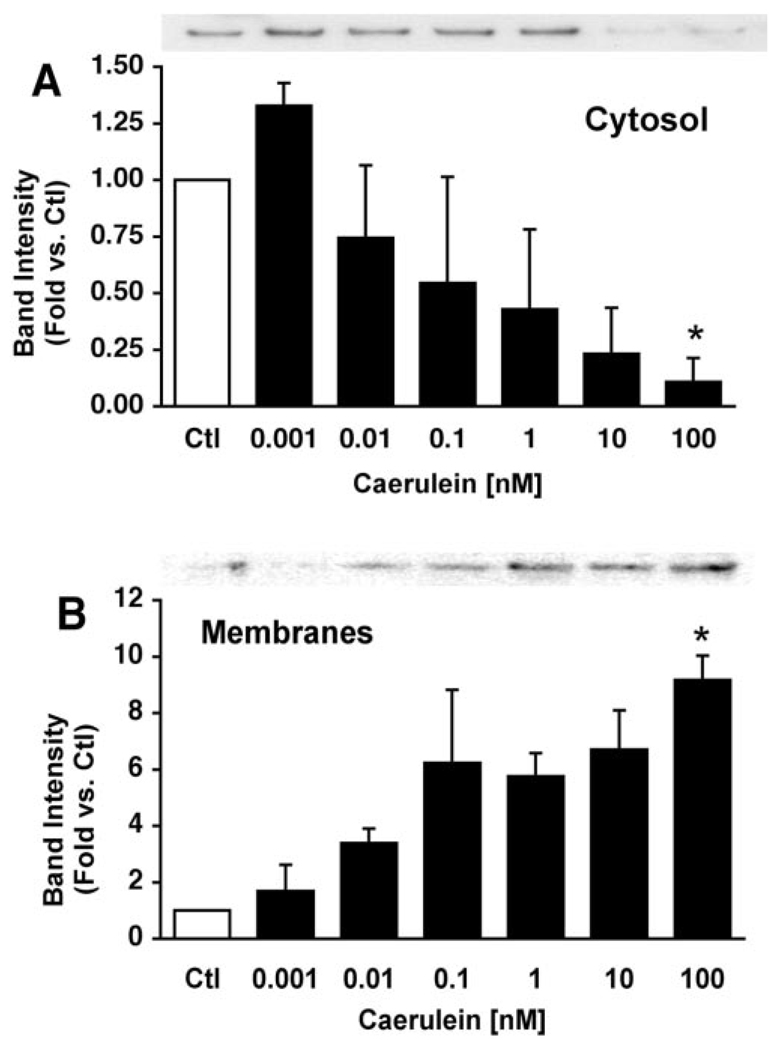
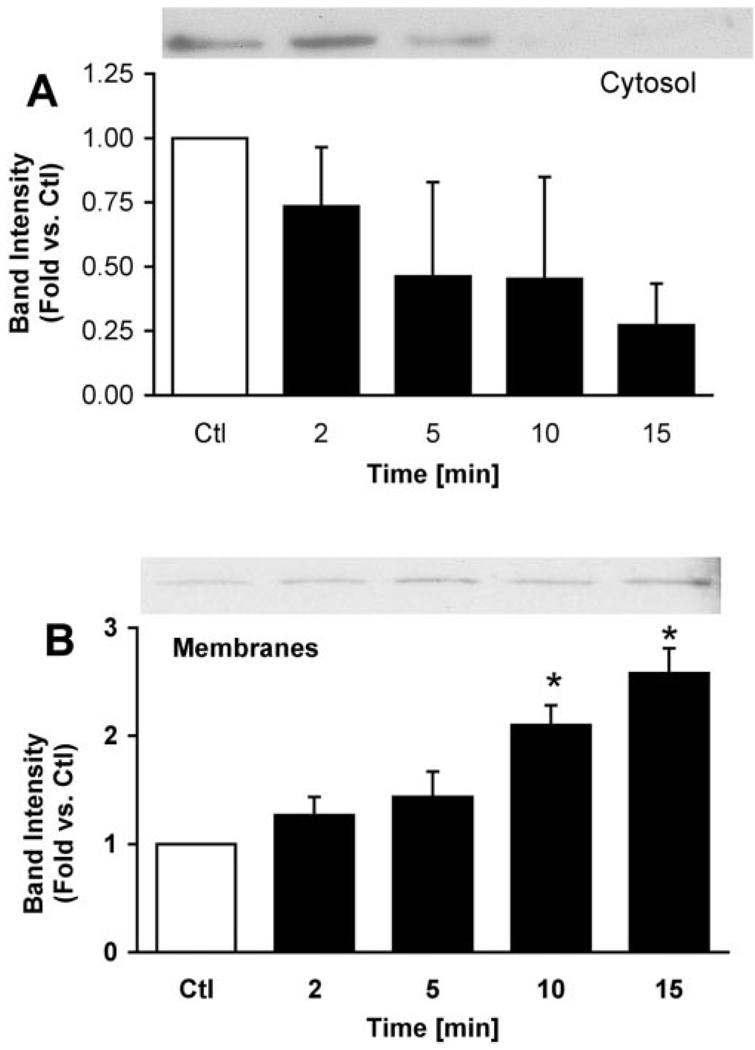
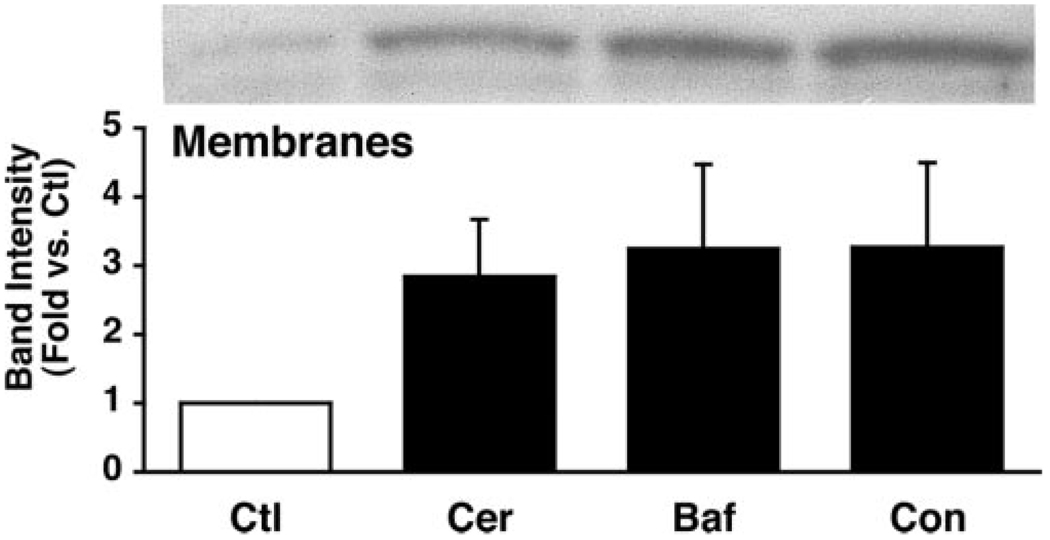
Supramaximal concentrations of cholecystokinin or its analogue caerulein have been shown to stimulate the proteolytic activation of zymogens within the pancreatic acinar cell and initiate acute pancreatitis. Previous studies suggest that a low pH compartment might be required for activation. To test this hypothesis, the effects of agents that modulate intracellular pH on caerulein-induced trypsin and chymotrypsin activation were studied. Pretreatment of pancreatic acini with the proto-ionophore monensin (10 microM) and the weak base chloroquine (40 microM) inhibited activation. Pre-incubation with the vacuolar ATPase (V-ATPase) inhibitors bafilomycin A(1) and concanamycin A also decreased activation in a concentration-dependent manner with 50% inhibition at approximately 50 and 25 nM, respectively. Caerulein stimulation caused a time- and concentration-dependent translocation of soluble V-ATPase V(1) subunits to a membrane fraction, a marker of V-ATPase activation. Carbachol also stimulated translocation at supramaximal concentrations. Elevation of cytosolic Ca(2+) by thapsigargin was sufficient to induce translocation. Thus, stimulation of V-ATPase activity appears to be required for agonist-induced zymogen activation in the pancreatic acinar cell.
Figures



References
-
- Saluja A, Saito I, Saluja M, Houlihan MJ, Powers RE, Meldolesi J, Steer M. Am. J. Physiol. 1985;249:G702–G710. - PubMed
-
- Leach SD, Bilchik AJ, Karapetian O, Gorelick FS, Modlin IM. Pancreas. 1993;8:64–69. - PubMed
-
- Guillaumes S, Blanco I, Villanueva A, Sans MD, Clave P, Chabas A, Farre A, Lluis F. Pancreas. 1997;14:262–266. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
