

Membrane trafficking and mitochondrial abnormalities precede subunit c deposition in a cerebellar cell model of juvenile neuronal ceroid lipofuscinosis
- PMID: 15588329
- PMCID: PMC539297
- DOI: 10.1186/1471-2202-5-57
Membrane trafficking and mitochondrial abnormalities precede subunit c deposition in a cerebellar cell model of juvenile neuronal ceroid lipofuscinosis
Abstract
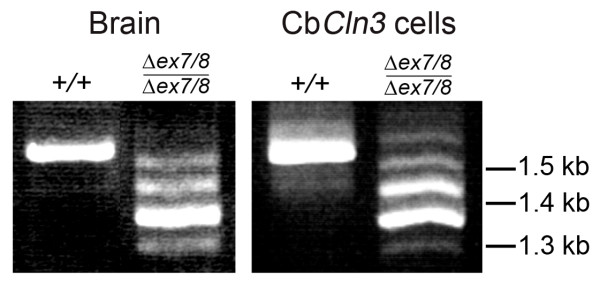
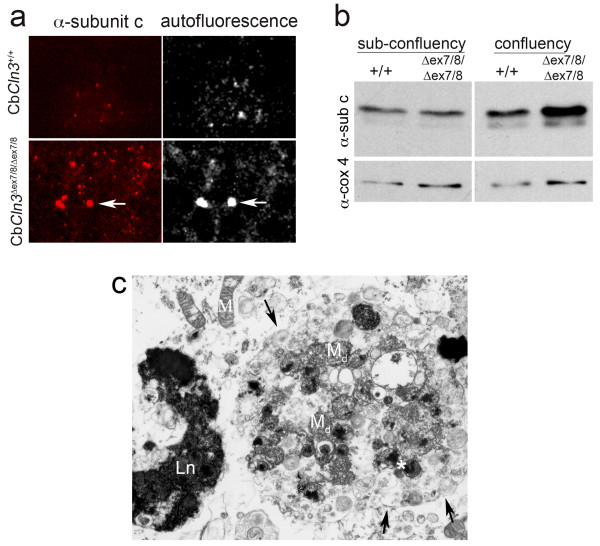
Background: JNCL is a recessively inherited, childhood-onset neurodegenerative disease most-commonly caused by a approximately 1 kb CLN3 mutation. The resulting loss of battenin activity leads to deposition of mitochondrial ATP synthase, subunit c and a specific loss of CNS neurons. We previously generated Cln3Deltaex7/8 knock-in mice, which replicate the common JNCL mutation, express mutant battenin and display JNCL-like pathology.
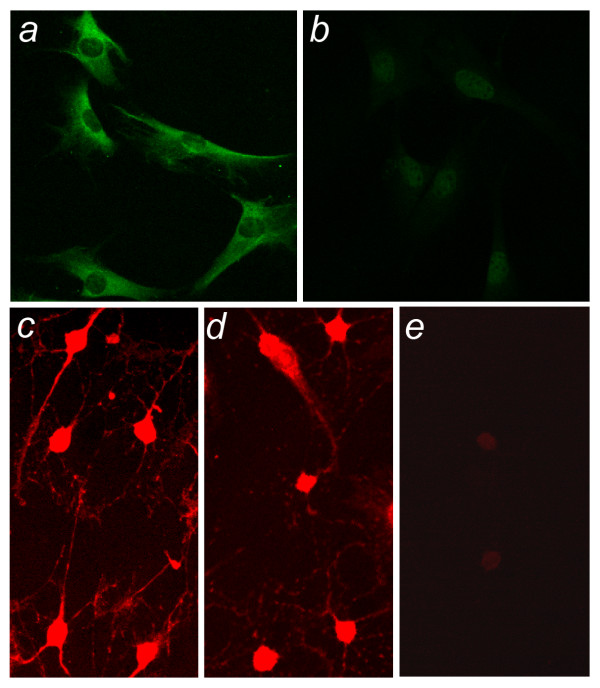
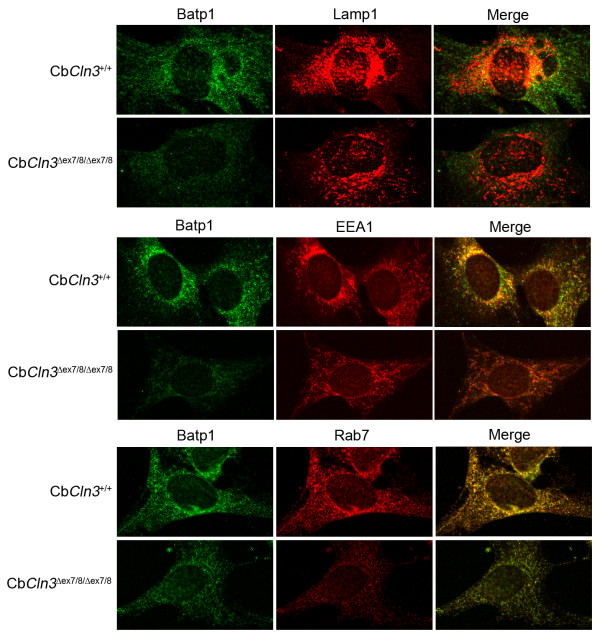
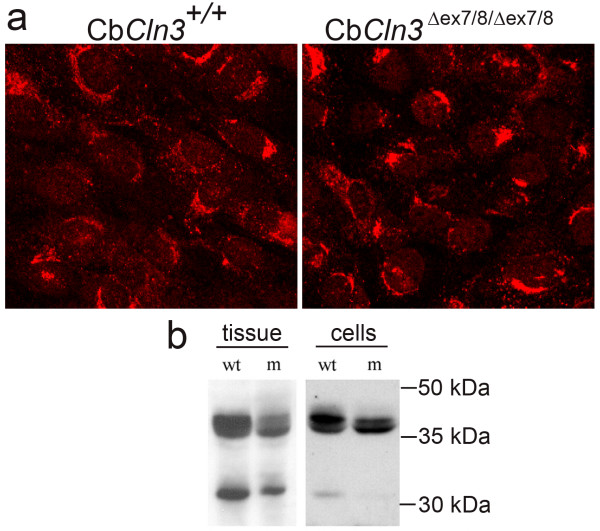
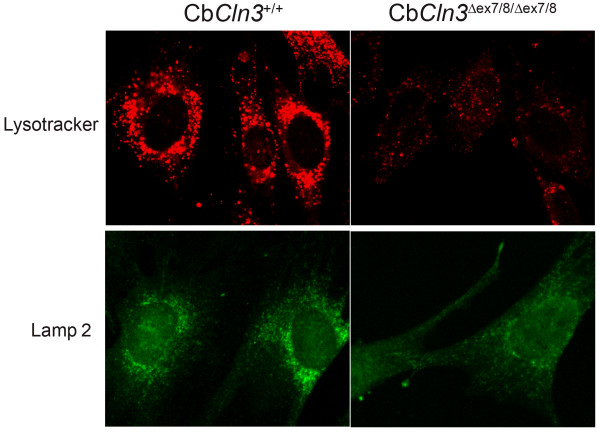
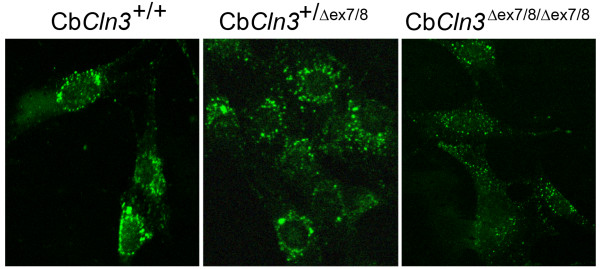
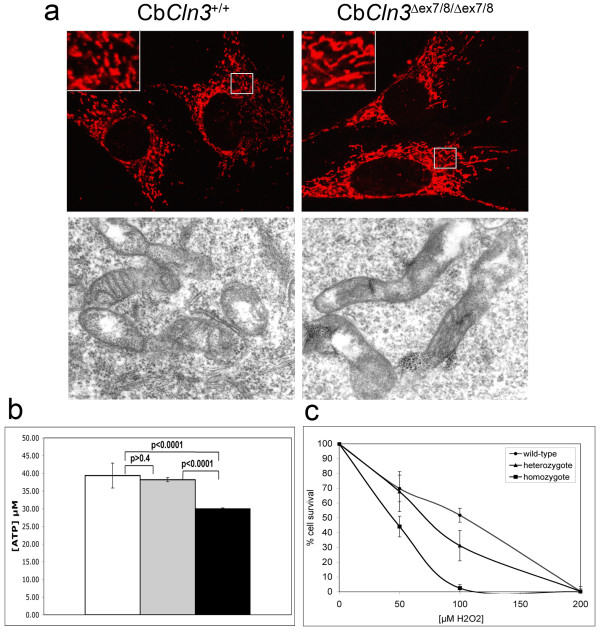
Results: To elucidate the consequences of the common JNCL mutation in neuronal cells, we used P4 knock-in mouse cerebella to establish conditionally immortalized CbCln3 wild-type, heterozygous, and homozygous neuronal precursor cell lines, which can be differentiated into MAP-2 and NeuN-positive, neuron-like cells. Homozygous CbCln3Deltaex7/8 precursor cells express low levels of mutant battenin and, when aged at confluency, accumulate ATPase subunit c. Recessive phenotypes are also observed at sub-confluent growth; cathepsin D transport and processing are altered, although enzyme activity is not significantly affected, lysosomal size and distribution are altered, and endocytosis is reduced. In addition, mitochondria are abnormally elongated, cellular ATP levels are decreased, and survival following oxidative stress is reduced.
Conclusions: These findings reveal that battenin is required for intracellular membrane trafficking and mitochondrial function. Moreover, these deficiencies are likely to be early events in the JNCL disease process and may particularly impact neuronal survival.
Figures
References
-
- Rider JA, Rider DL. Batten disease: past, present, and future. Am J Med Genet Suppl. 1988;5:21–26. - PubMed
-
- Jolly RD, Martinus RD, Palmer DN. Sheep and other animals with ceroid-lipofuscinoses: their relevance to Batten disease. Am J Med Genet. 1992;42:609–614. - PubMed
-
- Palmer DN, Fearnley IM, Walker JE, Hall NA, Lake BD, Wolfe LS, Haltia M, Martinus RD, Jolly RD. Mitochondrial ATP synthase subunit c storage in the ceroid-lipofuscinoses (Batten disease) Am J Med Genet. 1992;42:561–567. - PubMed
-
- Wisniewski KE, Rapin I, Heaney-Kieras J. Clinico-pathological variability in the childhood neuronal ceroid-lipofuscinoses and new observations on glycoprotein abnormalities. American Journal of Medical Genetics - Supplement. 1988;5:27–46. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
