

Molecular analysis of sister chromatid recombination in mammalian cells
- PMID: 15590323
- PMCID: PMC2967438
- DOI: 10.1016/j.dnarep.2004.08.010
Molecular analysis of sister chromatid recombination in mammalian cells
Abstract
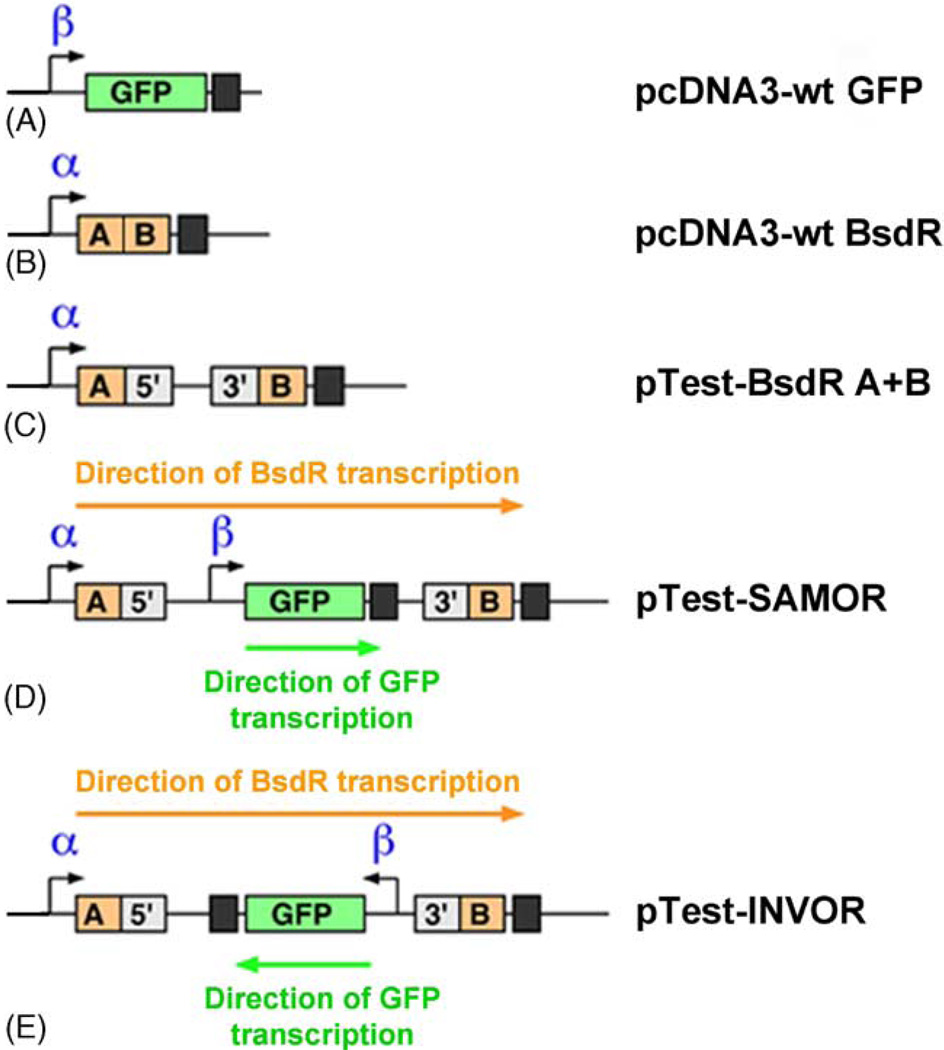
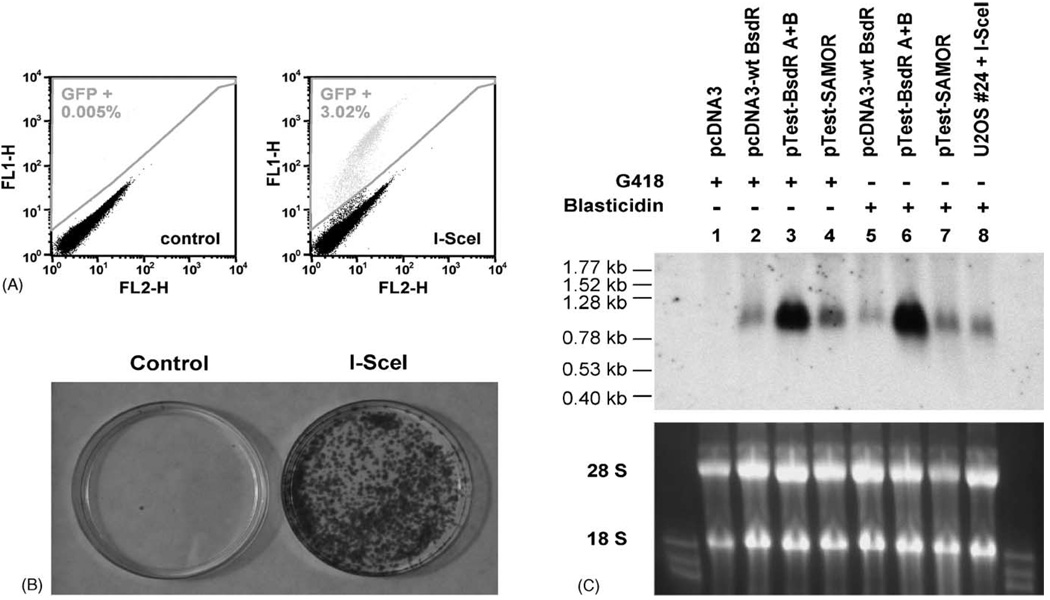
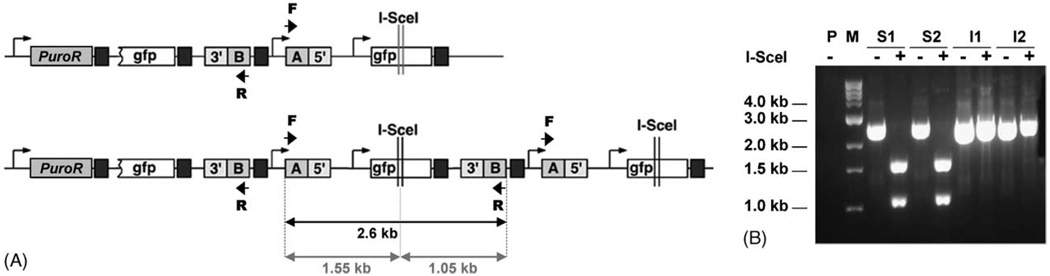
Sister chromatid recombination (SCR) is a potentially error-free pathway for the repair of double-strand breaks arising during replication and is thought to be important for the prevention of genomic instability and cancer. Analysis of sister chromatid recombination at a molecular level has been limited by the difficulty of selecting specifically for these events. To overcome this, we have developed a novel "nested intron" reporter that allows the positive selection in mammalian cells of "long tract" gene conversion events arising between sister chromatids. We show that these events arise spontaneously in cycling cells and are strongly induced by a site-specific double-strand break (DSB) caused by the restriction endonuclease, I-SceI. Notably, some I-SceI-induced sister chromatid recombination events entailed multiple rounds of gene amplification within the reporter, with the generation of a concatemer of amplified gene segments. Thus, there is an intimate relationship between sister chromatid recombination control and certain types of gene amplification. Dysregulated sister chromatid recombination may contribute to cancer progression, in part, by promoting gene amplification.
Figures



References
-
- Gonzalez-Barrera S, Cortes-Ledesma F, Wellinger RE, Aguilera A. Equal sister chromatid exchange is a major mechanism of double-strand break repair in yeast. Mol. Cell. 2003;11:1661–1671. - PubMed
-
- Cox MM. The non-mutagenic repair of broken replication forks via recombination. Mut. Res. 2002;510:107–120. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
