

Evolution of the Beckwith-Wiedemann syndrome region in vertebrates
- PMID: 15590939
- PMCID: PMC540281
- DOI: 10.1101/gr.2689805
Evolution of the Beckwith-Wiedemann syndrome region in vertebrates
Abstract
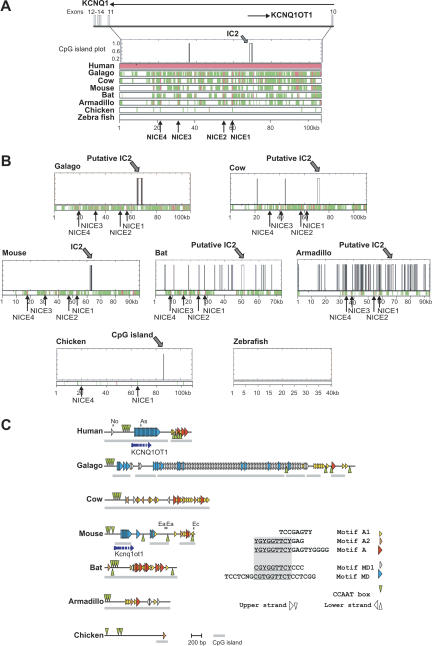
In the animal kingdom, genomic imprinting appears to be restricted to mammals. It remains an open question how structural features for imprinting evolved in mammalian genomes. The clustering of genes around imprinting control centers (ICs) is regarded as a hallmark for the coordinated imprinted regulation. Hence imprinted clusters might be structurally distinct between mammals and nonimprinted vertebrates. To address this question we compared the organization of the Beckwith Wiedemann syndrome (BWS) gene cluster in mammals, chicken, Fugu (pufferfish), and zebrafish. Our analysis shows that gene synteny is apparently well conserved between mammals and birds, and is detectable but less pronounced in fish. Hence, clustering apparently evolved during vertebrate radiation and involved two major duplication events that took place before the separation of the fish and mammalian lineages. A cross-species analysis of imprinting center regions showed that some structural features can already be recognized in nonimprinted amniotes in one of the imprinting centers (IC2). In contrast, the imprinting center IC1 is absent in chicken. This suggests a progressive and stepwise evolution of imprinting control elements. In line with that, imprinting centers in mammals apparently exhibit a high degree of structural and sequence variation despite conserved epigenetic marking.
Figures


Similar articles
-
Conserved synteny of mammalian imprinted genes in chicken, frog, and fish genomes.Cytogenet Genome Res. 2007;117(1-4):78-85. doi: 10.1159/000103167. Cytogenet Genome Res. 2007. PMID: 17675847
-
Molecular subtypes and phenotypic expression of Beckwith-Wiedemann syndrome.Eur J Hum Genet. 2005 Sep;13(9):1025-32. doi: 10.1038/sj.ejhg.5201463. Eur J Hum Genet. 2005. PMID: 15999116
-
Sequence and functional comparison in the Beckwith-Wiedemann region: implications for a novel imprinting centre and extended imprinting.Hum Mol Genet. 2000 Nov 1;9(18):2691-706. doi: 10.1093/hmg/9.18.2691. Hum Mol Genet. 2000. PMID: 11063728
-
Parent-of-origin specific QTL--a possibility towards understanding reciprocal effects in chicken and the origin of imprinting.Cytogenet Genome Res. 2007;117(1-4):305-12. doi: 10.1159/000103192. Cytogenet Genome Res. 2007. PMID: 17675872 Review.
-
Construction and evolution of imprinted loci in mammals.Trends Genet. 2007 Sep;23(9):440-8. doi: 10.1016/j.tig.2007.07.003. Epub 2007 Aug 1. Trends Genet. 2007. PMID: 17683825 Review.
Cited by
-
Kcnq1ot1/Lit1 noncoding RNA mediates transcriptional silencing by targeting to the perinucleolar region.Mol Cell Biol. 2008 Jun;28(11):3713-28. doi: 10.1128/MCB.02263-07. Epub 2008 Feb 25. Mol Cell Biol. 2008. PMID: 18299392 Free PMC article.
-
Genetic basis and identification of candidate genes for wooden breast and white striping in commercial broiler chickens.Sci Rep. 2021 Mar 24;11(1):6785. doi: 10.1038/s41598-021-86176-4. Sci Rep. 2021. PMID: 33762630 Free PMC article.
-
Imprinted genes show unique patterns of sequence conservation.BMC Genomics. 2010 Nov 22;11:649. doi: 10.1186/1471-2164-11-649. BMC Genomics. 2010. PMID: 21092170 Free PMC article.
-
Targeted gene suppression by inducing de novo DNA methylation in the gene promoter.Epigenetics Chromatin. 2014 Aug 18;7:20. doi: 10.1186/1756-8935-7-20. eCollection 2014. Epigenetics Chromatin. 2014. PMID: 25184003 Free PMC article.
-
Is imprinting in printing or in press?J Mol Med (Berl). 2005 May;83(5):321-3. doi: 10.1007/s00109-005-0653-5. J Mol Med (Berl). 2005. PMID: 15838693 No abstract available.
References
-
- Aparicio, S., Chapman, J., Stupka, E., Putnam, N., Chia, J.M., Dehal, P., Christoffels, A., Rash, S., Hoon, S., Smit, A. et al. 2002. Whole-genome shotgun assembly and analysis of the genome of Fugu rubripes. Science 297: 1301-1310. - PubMed
-
- Bailey, T.L. and Elkan, C. 1994. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 2: 28-36. - PubMed
-
- Bell, A.C. and Felsenfeld, G. 2000. Methylation of a CTCF-dependent boundary controls imprinted expression of the Igf2 gene. Nature 405: 482-485. - PubMed
Web site references
-
- http://www.ncbi.nlm.nih.gov; National Center for Biotechnology Information.
-
- http://www.ensembl.org/Fugu_rubripes/; Sanger Institute Fugu Genome Browser.
-
- http://www.ensembl.org; Sanger Institute Ensembl Genome Browser.
-
- http://cancer.otago.ac.nz/IGC/Web/home.html; Imprinted Gene Catalogue, University of Otago.
-
- http://www.mgu.har.mrc.ac.uk/imprinting/imprinting.html; Mammalian Genetics Unit, Harwell UK.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases