

Molecular origins of rapid and continuous morphological evolution
- PMID: 15596718
- PMCID: PMC539791
- DOI: 10.1073/pnas.0408118101
Molecular origins of rapid and continuous morphological evolution
Abstract
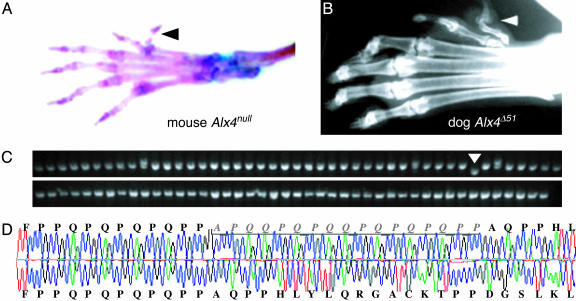
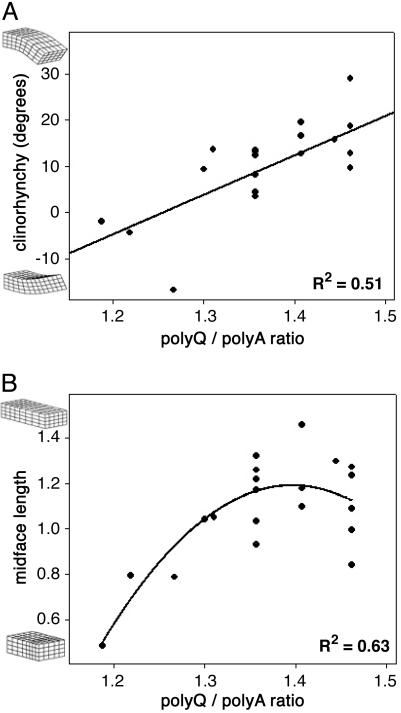
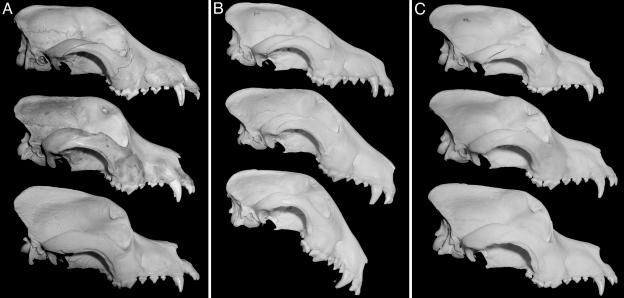
Mutations in cis-regulatory sequences have been implicated as being the predominant source of variation in morphological evolution. We offer a hypothesis that gene-associated tandem repeat expansions and contractions are a major source of phenotypic variation in evolution. Here, we describe a comparative genomic study of repetitive elements in developmental genes of 92 breeds of dogs. We find evidence for selection for divergence at coding repeat loci in the form of both elevated purity and extensive length polymorphism among different breeds. Variations in the number of repeats in the coding regions of the Alx-4 (aristaless-like 4) and Runx-2 (runt-related transcription factor 2) genes were quantitatively associated with significant differences in limb and skull morphology. We identified similar repeat length variation in the coding repeats of Runx-2, Twist, and Dlx-2 in several other species. The high frequency and incremental effects of repeat length mutations provide molecular explanations for swift, yet topologically conservative morphological evolution.
Figures
References
-
- Mayr, E. (1954) in Evolution as a Process, ed. Huxley, J. (Allen and Unwin, London), pp. 157–180.
-
- Eldredge, N. & Gould, S. J. (1972) in Models in Paleobiology, ed. Schopf, T. J. M. (Freeman & Cooper, San Francisco), pp. 305–332.
-
- Thompson, D'A. W. (1917) On Growth and Form (Dover, New York).
-
- Carroll, S. B. (2000) Cell 101, 577–580. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
