

Construction and maintenance of the optimal photosynthetic systems of the leaf, herbaceous plant and tree: an eco-developmental treatise
- PMID: 15598701
- PMCID: PMC4246796
- DOI: 10.1093/aob/mci049
Construction and maintenance of the optimal photosynthetic systems of the leaf, herbaceous plant and tree: an eco-developmental treatise
Abstract
Background and aims: The paper by Monsi and Saeki in 1953 (Japanese Journal of Botany 14: 22-52) was pioneering not only in mathematical modelling of canopy photosynthesis but also in eco-developmental studies of seasonal changes in leaf canopies.
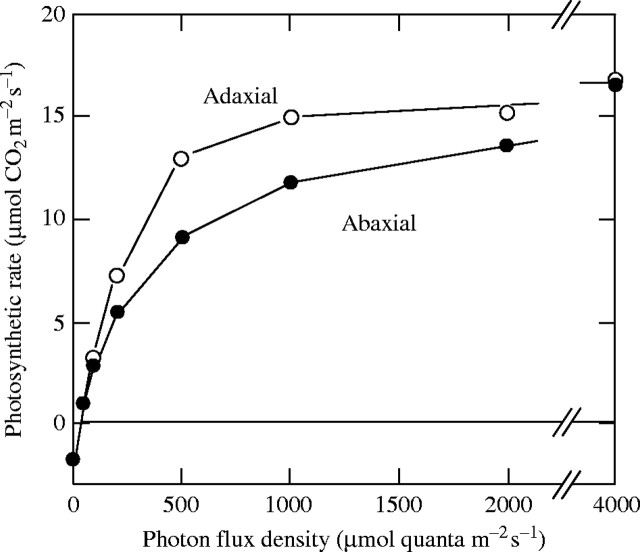
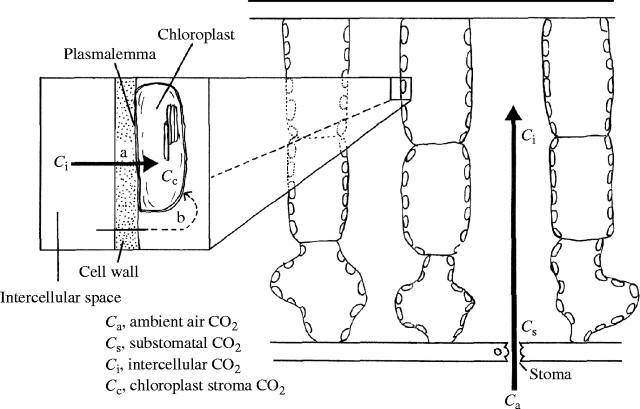
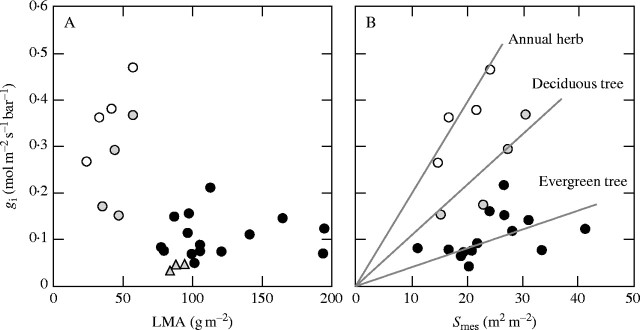
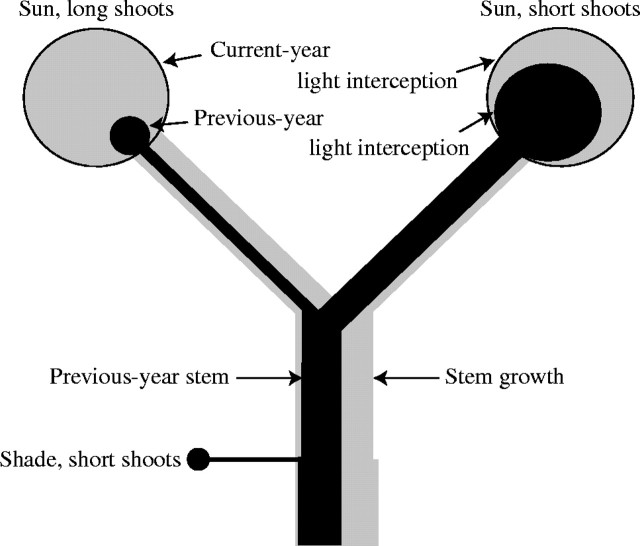
Scope: Construction and maintenance mechanisms of efficient photosynthetic systems at three different scaling levels--single leaves, herbaceous plants and trees--are reviewed mainly based on the nitrogen optimization theory. First, the nitrogen optimization theory with respect to the canopy and the single leaf is briefly introduced. Secondly, significance of leaf thickness in CO2 diffusion in the leaf and in leaf photosynthesis is discussed. Thirdly, mechanisms of adjustment of photosynthetic properties of the leaf within the herbaceous plant individual throughout its life are discussed. In particular, roles of sugar sensing, redox control and of cytokinin are highlighted. Finally, the development of a tree is considered.
Conclusions: Various mechanisms contribute to construction and maintenance of efficient photosynthetic systems. Molecular backgrounds of these ecologically important mechanisms should be clarified. The construction mechanisms of the tree cannot be explained solely by the nitrogen optimization theory. It is proposed that the pipe model theory in its differential form could be a potential tool in future studies in this research area.
Figures

Similar articles
-
Development of the Monsi-Saeki theory on canopy structure and function.Ann Bot. 2005 Feb;95(3):483-94. doi: 10.1093/aob/mci047. Epub 2004 Dec 7. Ann Bot. 2005. PMID: 15585544 Free PMC article.
-
Structure and function of plant canopies.Ann Bot. 2005 Feb;95(3):481-2. doi: 10.1093/aob/mci046. Epub 2004 Dec 7. Ann Bot. 2005. PMID: 15661750 Free PMC article.
-
Optimal photosynthetic use of light by tropical tree crowns achieved by adjustment of individual leaf angles and nitrogen content.Ann Bot. 2009 Mar;103(5):795-805. doi: 10.1093/aob/mcn265. Epub 2009 Jan 16. Ann Bot. 2009. PMID: 19151040 Free PMC article.
-
Leaf canopy as a dynamic system: ecophysiology and optimality in leaf turnover.Ann Bot. 2005 Feb;95(3):521-33. doi: 10.1093/aob/mci050. Epub 2004 Dec 7. Ann Bot. 2005. PMID: 15585542 Free PMC article. Review.
-
Variation in leaf photosynthetic capacity within plant canopies: optimization, structural, and physiological constraints and inefficiencies.Photosynth Res. 2023 Nov;158(2):131-149. doi: 10.1007/s11120-023-01043-9. Epub 2023 Aug 24. Photosynth Res. 2023. PMID: 37615905 Review.
Cited by
-
Supplemental Upward Lighting from Underneath to Obtain Higher Marketable Lettuce (Lactuca sativa) Leaf Fresh Weight by Retarding Senescence of Outer Leaves.Front Plant Sci. 2015 Dec 14;6:1110. doi: 10.3389/fpls.2015.01110. eCollection 2015. Front Plant Sci. 2015. PMID: 26697055 Free PMC article.
-
Anatomical acclimation of mature leaves to increased irradiance in sycamore maple (Acer pseudoplatanus L.).Photosynth Res. 2022 Oct;154(1):41-55. doi: 10.1007/s11120-022-00953-4. Epub 2022 Sep 3. Photosynth Res. 2022. PMID: 36057003 Free PMC article.
-
If self-shading is so bad, why is there so much? Short shoots reconcile costs and benefits.New Phytol. 2023 Mar;237(5):1684-1695. doi: 10.1111/nph.18636. Epub 2022 Dec 21. New Phytol. 2023. PMID: 36427292 Free PMC article.
-
Cytokinin import rate as a signal for photosynthetic acclimation to canopy light gradients.Plant Physiol. 2007 Apr;143(4):1841-52. doi: 10.1104/pp.106.094631. Epub 2007 Feb 2. Plant Physiol. 2007. PMID: 17277095 Free PMC article.
-
Fagaceae tree species allocate higher fraction of nitrogen to photosynthetic apparatus than Leguminosae in Jianfengling tropical montane rain forest, China.PLoS One. 2018 Feb 1;13(2):e0192040. doi: 10.1371/journal.pone.0192040. eCollection 2018. PLoS One. 2018. PMID: 29390007 Free PMC article.
References
-
- Ackerly DD. 1992. Light, leaf age and nitrogen concentration in a tropical vine. Oecologia 89: 596–600. - PubMed
-
- Anderson JM. 1986. Photoregulation of the composition, function, and structure of thylakoid membranes. Annual Review of Plant Physiology 37: 93–136.
-
- Anten NPR, Hikosaka K, Hirose T. 2000. Nitrogen utilisation and the photosynthetic system. In: Marshall B, Roberts JA, eds. Leaf development and canopy growth. Sheffield: Sheffield Academic Press, 171–203.
-
- Björkman O. 1981. Responses to different quantum flux densities. In: Lange OL, Nobel PS, Osmond CB, Ziegler H, eds. Physiological plant ecology. I. Responses to the physical environment. Berlin: Springer-Verlag, 57–107.
