

Activation of CRABS CLAW in the Nectaries and Carpels of Arabidopsis
- PMID: 15598802
- PMCID: PMC544487
- DOI: 10.1105/tpc.104.026666
Activation of CRABS CLAW in the Nectaries and Carpels of Arabidopsis
Abstract
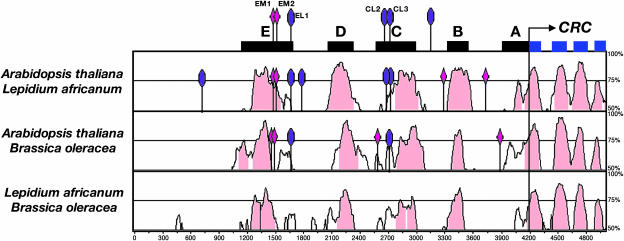
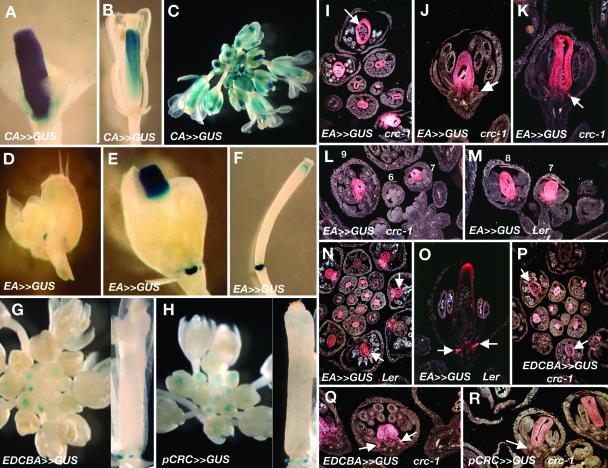
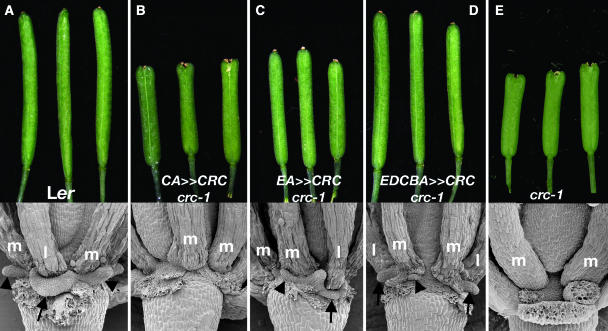
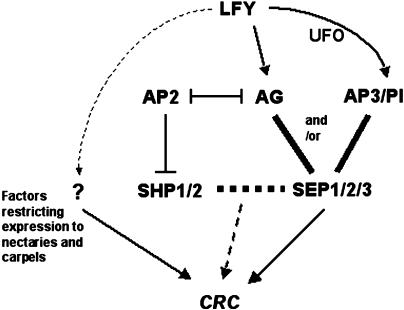
CRABS CLAW (CRC), a member of the YABBY gene family, is required for nectary and carpel development. To further understand CRC regulation in Arabidopsis thaliana, we performed phylogenetic footprinting analyses of 5' upstream regions of CRC orthologs from three Brassicaceae species, including Arabidopsis. Phylogenetic footprinting efficiently identified functionally important regulatory regions (modules), indicating that CRC expression is regulated by a combination of positive and negative regulatory elements in the modules. Within the conserved modules, we identified putative binding sites of LEAFY and MADS box proteins, and functional in vivo analyses revealed their importance for CRC expression. Both expression and genetic studies demonstrate that potential binding sites for MADS box proteins within the conserved regions are functionally significant for the transcriptional regulation of CRC in nectaries. We propose that in wild-type flowers, a combination of floral homeotic gene activities, specifically the B class genes APETALA3 and PISTILLATA and the C class gene AGAMOUS act redundantly with each other and in combination with SEPALLATA genes to activate CRC in the nectaries and carpels. In the absence of B and C class gene activities, other genes such as SHATTERPROOF1/2 can substitute if they are ectopically expressed, as in an A class mutant background (apetala2). These MADS box proteins may provide general floral factors that must work in conjunction with specific factors in the activation of CRC in the nectaries and carpels.
Figures

Similar articles
-
CRABS CLAW, a gene that regulates carpel and nectary development in Arabidopsis, encodes a novel protein with zinc finger and helix-loop-helix domains.Development. 1999 Jun;126(11):2387-96. doi: 10.1242/dev.126.11.2387. Development. 1999. PMID: 10225998
-
The Floral C-Lineage Genes Trigger Nectary Development in Petunia and Arabidopsis.Plant Cell. 2018 Sep;30(9):2020-2037. doi: 10.1105/tpc.18.00425. Epub 2018 Aug 7. Plant Cell. 2018. PMID: 30087206 Free PMC article.
-
Regulatory elements of the floral homeotic gene AGAMOUS identified by phylogenetic footprinting and shadowing.Plant Cell. 2003 Jun;15(6):1296-309. doi: 10.1105/tpc.009548. Plant Cell. 2003. PMID: 12782724 Free PMC article.
-
Gene duplication and the evolution of plant MADS-box transcription factors.J Genet Genomics. 2012 Apr 20;39(4):157-65. doi: 10.1016/j.jgg.2012.02.008. Epub 2012 Mar 17. J Genet Genomics. 2012. PMID: 22546537 Review.
-
The autonomous pathway: epigenetic and post-transcriptional gene regulation in the control of Arabidopsis flowering time.Curr Opin Plant Biol. 2004 Oct;7(5):570-4. doi: 10.1016/j.pbi.2004.07.002. Curr Opin Plant Biol. 2004. PMID: 15337100 Review.
Cited by
-
Diversity and evolution of a trait mediating ant-plant interactions: insights from extrafloral nectaries in Senna (Leguminosae).Ann Bot. 2013 Jun;111(6):1263-75. doi: 10.1093/aob/mcs226. Epub 2012 Oct 26. Ann Bot. 2013. PMID: 23104672 Free PMC article.
-
The Roles of Plant Hormones and Their Interactions with Regulatory Genes in Determining Meristem Activity.Int J Mol Sci. 2019 Aug 20;20(16):4065. doi: 10.3390/ijms20164065. Int J Mol Sci. 2019. PMID: 31434317 Free PMC article. Review.
-
The evolution of floral nectaries in Disa (Orchidaceae: Disinae): recapitulation or diversifying innovation?Ann Bot. 2013 Nov;112(7):1303-19. doi: 10.1093/aob/mct197. Epub 2013 Aug 29. Ann Bot. 2013. PMID: 23997231 Free PMC article.
-
BELL1 interacts with CRABS CLAW and INNER NO OUTER to regulate ovule and seed development in pomegranate.Plant Physiol. 2023 Feb 12;191(2):1066-1083. doi: 10.1093/plphys/kiac554. Plant Physiol. 2023. PMID: 36477345 Free PMC article.
-
Nectary development in Cleome violacea.Front Plant Sci. 2023 Feb 9;13:1085900. doi: 10.3389/fpls.2022.1085900. eCollection 2022. Front Plant Sci. 2023. PMID: 36844906 Free PMC article.
References
-
- Alvarez, J., and Smyth, D.R. (1999). CRABS CLAW and SPATULA, two Arabidopsis genes that control carpel development in parallel with AGAMOUS. Development 126, 2377–2386. - PubMed
-
- Arnone, M.I., and Davidson, E.H. (1997). The hardwiring of development: Organization and function of genomic regulatory systems. Development 124, 1851–1864. - PubMed
-
- Baum, S.F., Eshed, Y., and Bowman, J.L. (2001). The Arabidopsis nectary is an ABC-independent floral structure. Development 128, 4657–4667. - PubMed
-
- Bowman, J.L., and Smyth, D.R. (1998). Patterns of petal and stamen reduction in Australian species of Lepidium L. (Brassicaceae). Int. J. Plant Sci. 159, 65–74.
-
- Bowman, J.L., and Smyth, D.R. (1999). CRABS CLAW, a gene that regulates carpel and nectary development in Arabidopsis, encodes a novel protein with zinc finger and helix-loop-helix domains. Development 126, 2387–2396. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
