

Regulation of uptake and processing of the quorum-sensing autoinducer AI-2 in Escherichia coli
- PMID: 15601708
- PMCID: PMC538819
- DOI: 10.1128/JB.187.1.238-248.2005
Regulation of uptake and processing of the quorum-sensing autoinducer AI-2 in Escherichia coli
Abstract
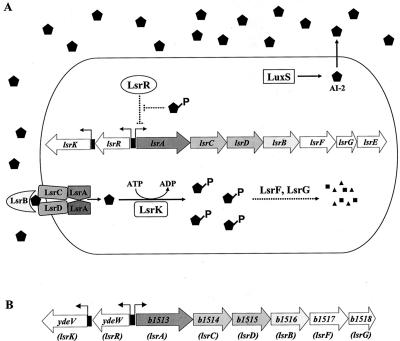
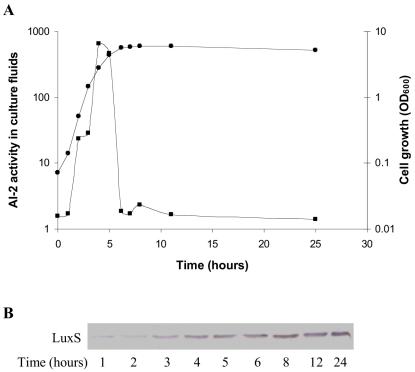
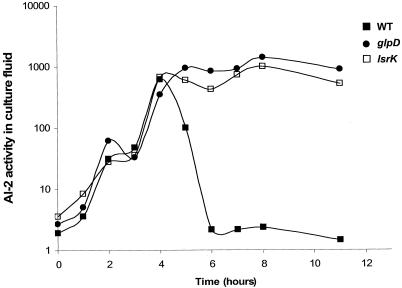
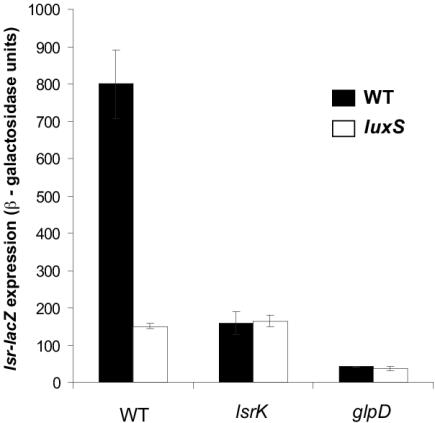
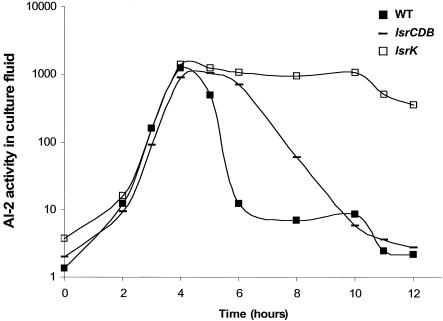
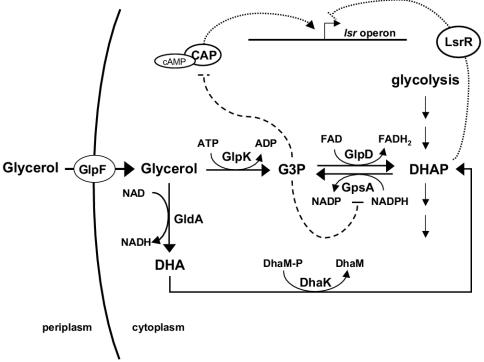
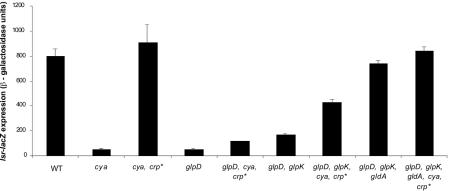
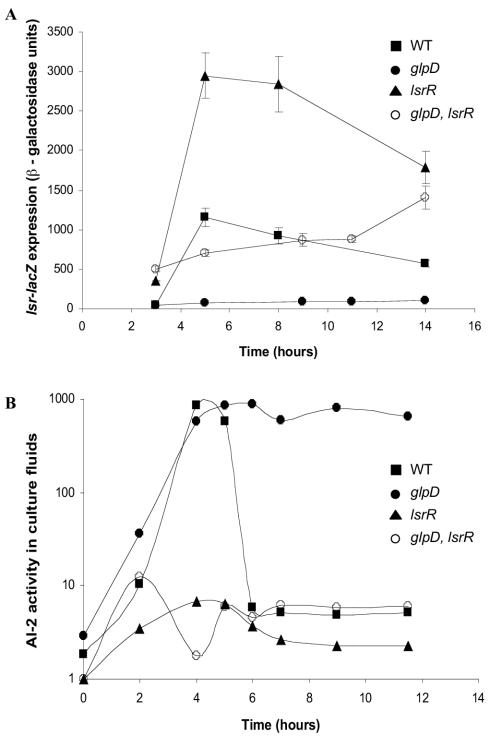
AI-2 is a quorum-sensing signaling molecule proposed to be involved in interspecies communication. In Escherichia coli and Salmonella enterica serovar Typhimurium, extracellular AI-2 accumulates in exponential phase, but the amount decreases drastically upon entry into stationary phase. In S. enterica serovar Typhimurium, the reduction in activity is due to import and processing of AI-2 by the Lsr transporter. We show that the Lsr transporter is functional in E. coli, and screening for mutants defective in AI-2 internalization revealed lsrK and glpD. Unlike the wild type, lsrK and glpD mutants do not activate transcription of the lsr operon in response to AI-2. lsrK encodes the AI-2 kinase, and the lsrK mutant fails to activate lsr expression because it cannot produce phospho-AI-2, which is the lsr operon inducer. glpD encodes the glycerol-3-phosphate (G3P) dehydrogenase, which is involved in glycerol and G3P metabolism. G3P accumulates in the glpD mutant and represses lsr transcription by preventing cyclic AMP (cAMP)-catabolite activator protein (CAP)-dependent activation. Dihydroxyacetone phosphate (DHAP) also accumulates in the glpD mutant, and DHAP represses lsr transcription by a cAMP-CAP-independent mechanism involving LsrR, the lsr operon repressor. The requirement for cAMP-CAP in lsr activation explains why AI-2 persists in culture fluids of bacteria grown in media containing sugars that cause catabolite repression. These findings show that, depending on the prevailing growth conditions, the amount of time that the AI-2 signal is present and, in turn, the time that a given community of bacteria remains exposed to this signal can vary greatly.
Figures
Similar articles
-
Cyclic AMP (cAMP) and cAMP receptor protein influence both synthesis and uptake of extracellular autoinducer 2 in Escherichia coli.J Bacteriol. 2005 Mar;187(6):2066-76. doi: 10.1128/JB.187.6.2066-2076.2005. J Bacteriol. 2005. PMID: 15743955 Free PMC article.
-
LsrR-binding site recognition and regulatory characteristics in Escherichia coli AI-2 quorum sensing.Cell Res. 2009 Nov;19(11):1258-68. doi: 10.1038/cr.2009.91. Epub 2009 Jul 28. Cell Res. 2009. PMID: 19636340
-
Lsr-mediated transport and processing of AI-2 in Salmonella typhimurium.Mol Microbiol. 2003 Nov;50(4):1411-27. doi: 10.1046/j.1365-2958.2003.03781.x. Mol Microbiol. 2003. PMID: 14622426
-
Cell-to-cell signalling in Escherichia coli and Salmonella enterica.Mol Microbiol. 2004 May;52(4):933-45. doi: 10.1111/j.1365-2958.2004.04054.x. Mol Microbiol. 2004. PMID: 15130116 Review.
-
Bacterial interspecies quorum sensing in the mammalian gut microbiota.C R Biol. 2018 May-Jun;341(5):297-299. doi: 10.1016/j.crvi.2018.03.006. Epub 2018 Apr 7. C R Biol. 2018. PMID: 29631889 Review.
Cited by
-
Thiophenone Attenuates Enteropathogenic Escherichia coli O103:H2 Virulence by Interfering with AI-2 Signaling.PLoS One. 2016 Jun 16;11(6):e0157334. doi: 10.1371/journal.pone.0157334. eCollection 2016. PLoS One. 2016. PMID: 27309855 Free PMC article.
-
Effect of inactivation of luxS gene on the properties of Serratia proteamaculans 94 strain.Folia Microbiol (Praha). 2019 May;64(3):265-272. doi: 10.1007/s12223-018-0657-5. Epub 2018 Oct 25. Folia Microbiol (Praha). 2019. PMID: 30361877
-
Quorum Sensing Modulates the Epibiotic-Parasitic Relationship Between Actinomyces odontolyticus and Its Saccharibacteria epibiont, a Nanosynbacter lyticus Strain, TM7x.Front Microbiol. 2018 Sep 24;9:2049. doi: 10.3389/fmicb.2018.02049. eCollection 2018. Front Microbiol. 2018. PMID: 30319555 Free PMC article.
-
Single mutation makes Escherichia coli an insect mutualist.Nat Microbiol. 2022 Aug;7(8):1141-1150. doi: 10.1038/s41564-022-01179-9. Epub 2022 Aug 4. Nat Microbiol. 2022. PMID: 35927448 Free PMC article.
-
Agent-Based Modeling Demonstrates How Local Chemotactic Behavior Can Shape Biofilm Architecture.mSphere. 2019 May 29;4(3):e00285-19. doi: 10.1128/mSphere.00285-19. mSphere. 2019. PMID: 31142622 Free PMC article.
References
-
- Bassler, B. L., M. Wright, R. E. Showalter, and M. R. Silverman. 1993. Intercellular signalling in Vibrio harveyi: sequence and function of genes regulating expression of luminescence. Mol. Microbiol. 9:773-786. - PubMed
-
- Bassler, B. L., M. Wright, and M. R. Silverman. 1994. Multiple signalling systems controlling expression of luminescence in Vibrio harveyi: sequence and function of genes encoding a second sensory pathway. Mol. Microbiol. 13:273-286. - PubMed
-
- Bell, R. M., and J. E. Cronan, Jr. 1975. Mutants of Escherichia coli defective in membrane phospholipid synthesis. Phenotypic suppression of sn-glycerol-3-phosphate acyltransferase Km mutants by loss of feedback inhibition of the biosynthetic sn-glycerol-3-phosphate dehydrogenase. J. Biol. Chem. 250:7153-7158. - PubMed
-
- Blattner, F. R., G. Plunkett III, C. A. Bloch, N. T. Perna, V. Burland, M. Riley, J. Collado-Vides, J. D. Glasner, C. K. Rode, G. F. Mayhew, J. Gregor, N. W. Davis, H. A. Kirkpatrick, M. A. Goeden, D. J. Rose, B. Mau, and Y. Shao. 1997. The complete genome sequence of Escherichia coli K-12. Science 277:1453-1474. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
