

Coordination of replication and transcription along a Drosophila chromosome
- PMID: 15601823
- PMCID: PMC535919
- DOI: 10.1101/gad.1246404
Coordination of replication and transcription along a Drosophila chromosome
Abstract
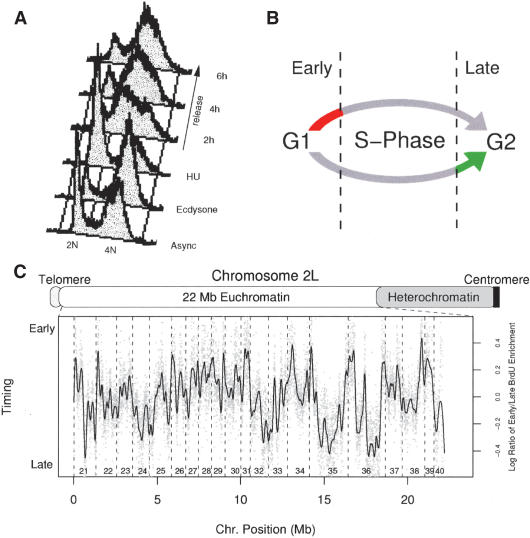
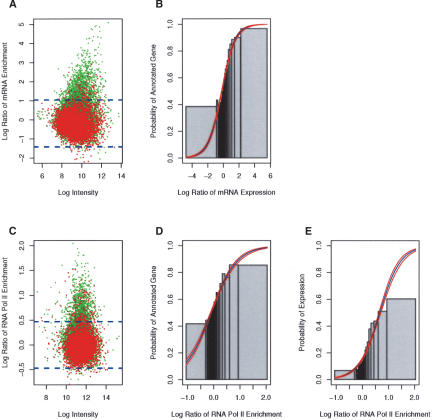
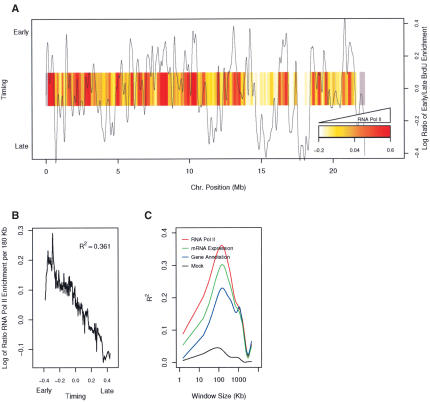
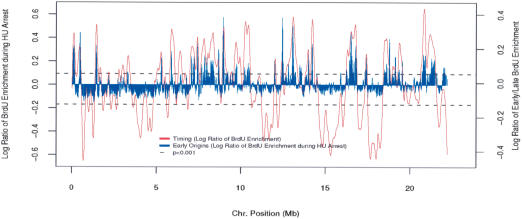
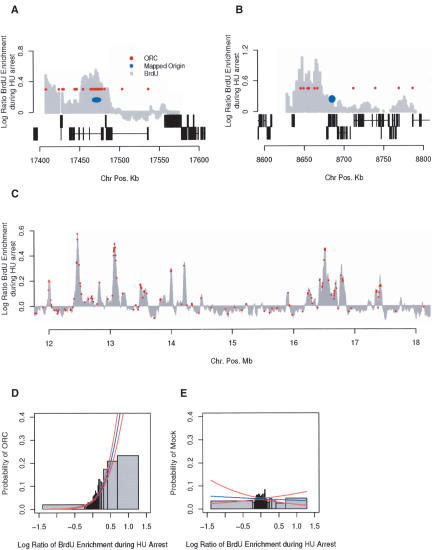
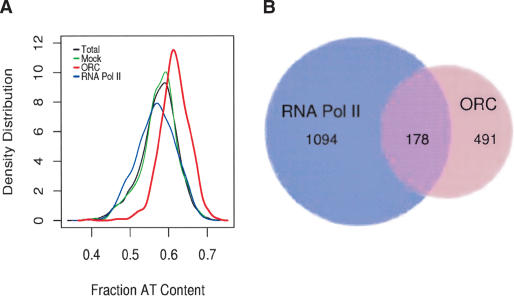
The mechanisms by which metazoan origins of DNA replication are defined, regulated, and influenced by chromosomal events remain poorly understood. To gain insights into these mechanisms, we developed a systematic approach using a Drosophila high-resolution genomic microarray to determine replication timing, identify replication origins, and map protein-binding sites along a chromosome arm. We identify a defined temporal pattern of replication that correlates with the density of active transcription. These data indicate that the influence of transcription status on replication timing is exerted over large domains (>100 kb) rather than at the level of individual genes. We identify 62 early activating replication origins across the chromosome by mapping sites of nucleotide incorporation during hydroxyurea arrest. Using genome-wide location analysis, we demonstrate that the origin recognition complex (ORC) is localized to specific chromosomal sites, many of which coincide with early activating origins. The molecular attributes of ORC-binding sites include increased AT-content and association with a subset of RNA Pol II-binding sites. Based on these findings, we suggest that the distribution of transcription along the chromosome acts locally to influence origin selection and globally to regulate origin activation.
Figures
References
-
- Abdurashidova G., Deganuto, M., Klima, R., Riva, S., Biamonti, G., Giacca, M., and Falaschi, A. 2000. Start sites of bidirectional DNA synthesis at the human lamin B2 origin. Science 287: 2023-2026. - PubMed
-
- Aladjem M., Groudine, M., Brody, L., Dieken, E., Fournier, R., Wahl, G., and Epner, E. 1995. Participation of the human β-globin locus control region in initiation of DNA replication. Science 270: 815-819. - PubMed
-
- Anglana M., Apiou, F., Bensimon, A., and Debatisse, M. 2003. Dynamics of DNA replication in mammalian somatic cells: Nucleotide pool modulates origin choice and interorigin spacing. Cell 114: 385-394. - PubMed
-
- Beall E., Manak, J., Zhou, S., Bell, M., Lipsick, J., and Botchan, M. 2002. Role for a Drosophila Myb-containing protein complex in site-specific DNA replication. Nature 420: 833-837. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases