

Molecular mechanism of hTid-1, the human homolog of Drosophila tumor suppressor l(2)Tid, in the regulation of NF-kappaB activity and suppression of tumor growth
- PMID: 15601829
- PMCID: PMC538758
- DOI: 10.1128/MCB.25.1.44-59.2005
Molecular mechanism of hTid-1, the human homolog of Drosophila tumor suppressor l(2)Tid, in the regulation of NF-kappaB activity and suppression of tumor growth
Abstract
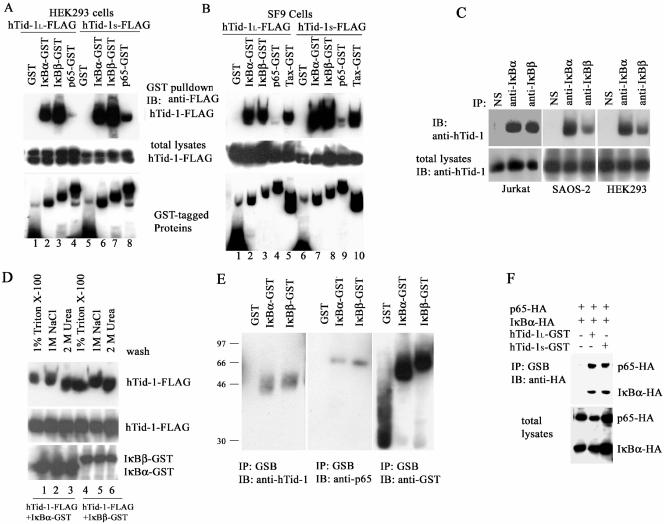
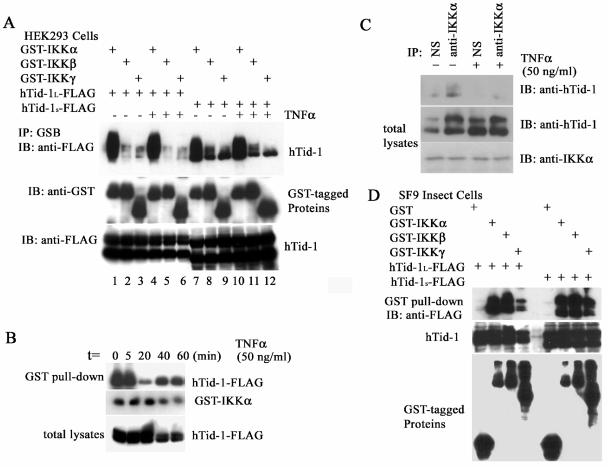
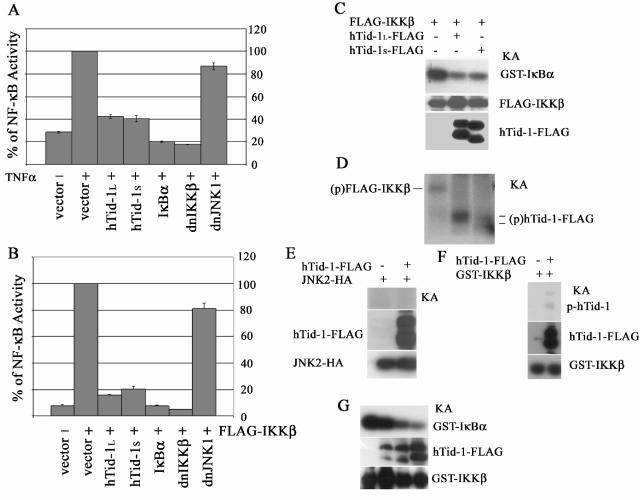
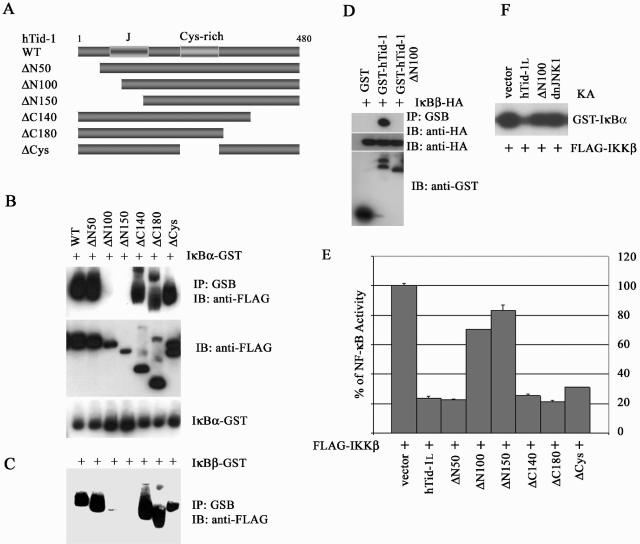
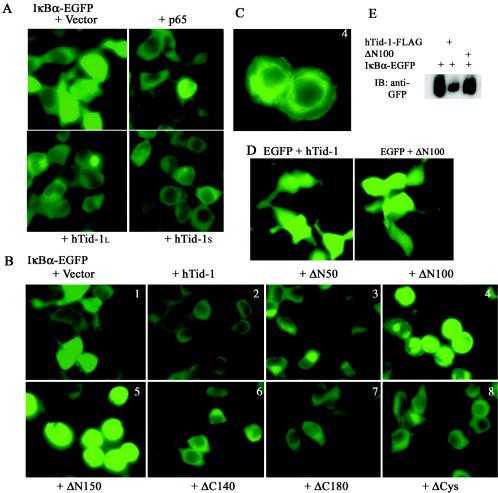
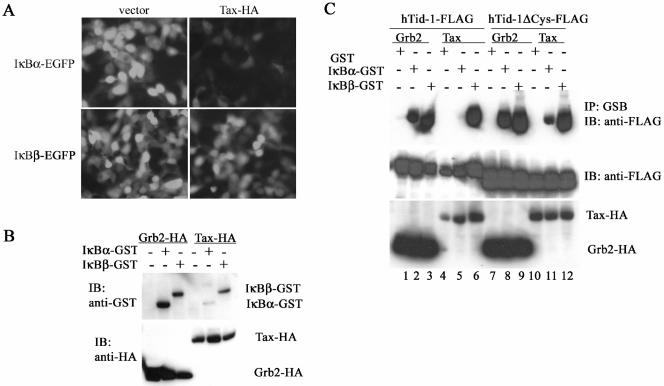
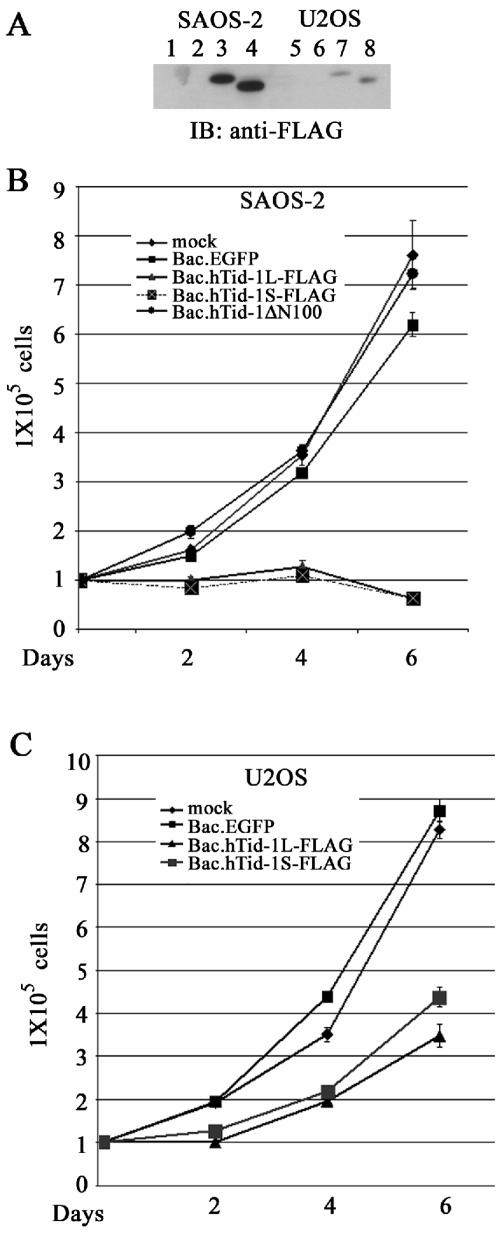
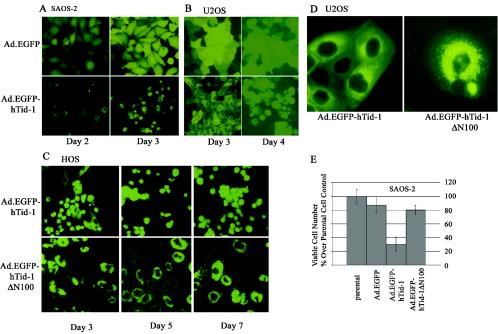
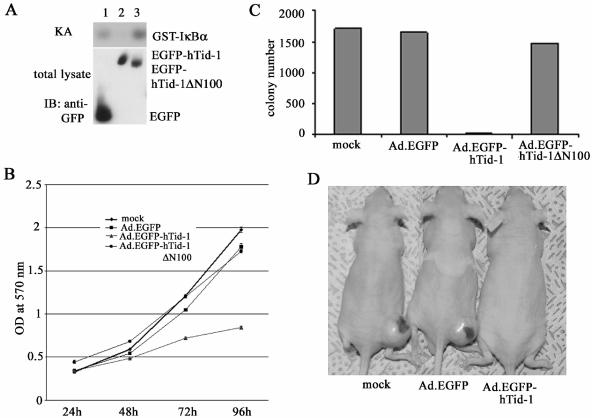
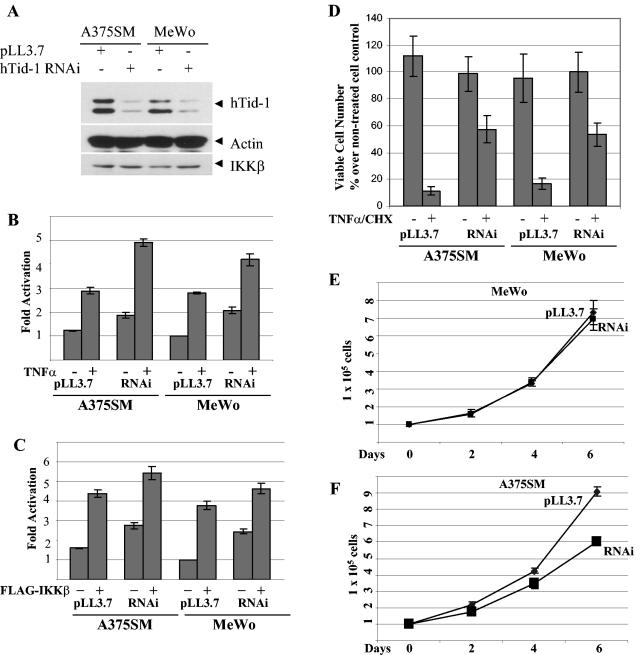
hTid-1, a human homolog of the Drosophila tumor suppressor l(2)Tid and a novel DnaJ protein, regulates the activity of nuclear factor kappaB (NF-kappaB), but its mechanism is not established. We report here that hTid-1 strongly associated with the cytoplasmic protein complex of NF-kappaB-IkappaB through direct interaction with IkappaBalpha/beta and the IKKalpha/beta subunits of the IkappaB kinase complex. These interactions resulted in suppression of the IKK activity in a J-domain-dependent fashion and led to the cytoplasmic retention and enhanced stability of IkappaB. Overexpression of hTid-1 by using recombinant baculovirus or adenovirus led to inhibition of cell proliferation and induction of apoptosis of human osteosarcoma cells regardless of the p53 expression status. Adherent cultured cells transduced with Ad.hTid-1 detached from the dish surface. Morphological changes consistent with apoptosis and cell death were evident 48 h after Ad.EGFP-hTid-1 transduction. In contrast, cells transduced with Ad.EGFP or Ad.EGFP-hTd-1DeltaN100, a mutant that has the N-terminal J domain deletion and that lost suppressive activity on IKK, continued to proliferate. Similar data were obtained with A375 human melanoma cells. Ad.EGFP or Ad.EGFP-hTd-1DeltaN100 ex vivo-transduced A375 cells injected subcutaneously into nude mice produced growing tumors, whereas Ad.EGFP-hTid-1-transduced cells did not. Collectively, the data suggest that hTid-1 represses the activity of NF-kappaB through physical and functional interactions with the IKK complex and IkappaB and, in doing so, it modulates cell growth and death.
Figures
References
-
- Agou, F., F. Ye, S. Goffinont, G. Courtois, S. Yamaoka, A. Israel, and M. Veron. 2002. NEMO trimerizes through its coiled-coil C-terminal domain. J. Biol. Chem. 277:17464-17475. - PubMed
-
- Arenzana-Seisdedos, F., P. Turpin, M. Rodriguez, D. Thomas, R. T. Hay, J. L. Virelizier, and C. Dargemont. 1997. Nuclear localization of IκBα promotes active transport of NF-κB from the nucleus to the cytoplasm. J. Cell Sci. 110:369-378. - PubMed
-
- Asea, A., M. Rehli, E. Kabingu, J. A. Boch, O. Bare, P. E. Auron, M. A. Stevenson, and S. K. Calderwood. 2002. Novel signal transduction pathway utilized by extracellular HSP70: role of Toll-like receptor (TLR) 2 and TLR4. J. Biol. Chem. 277:15028-15034. - PubMed
-
- Bakker, T. R., D. Reed, T. Renno, and C. V. Jongeneel. 1999. Efficient adenoviral transfer of NF-κB inhibitor sensitizes melanoma to tumor necrosis factor-mediated apoptosis. Int. J. Cancer 80:320-323. - PubMed
-
- Canamasas, I., A. Debes, P. G. Natali, and U. Kurzik-Dumke. 2003. Understanding human cancer using Drosophila: Tid47, a cytosolic product of the DnaJ-like tumor suppressor gene l2Tid, is a novel molecular partner of patched related to skin cancer. J. Biol. Chem. 278:30952-309060. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous