

Overcoming functional redundancy to elicit pachyonychia congenita-like nail lesions in transgenic mice
- PMID: 15601842
- PMCID: PMC538767
- DOI: 10.1128/MCB.25.1.197-205.2005
Overcoming functional redundancy to elicit pachyonychia congenita-like nail lesions in transgenic mice
Abstract
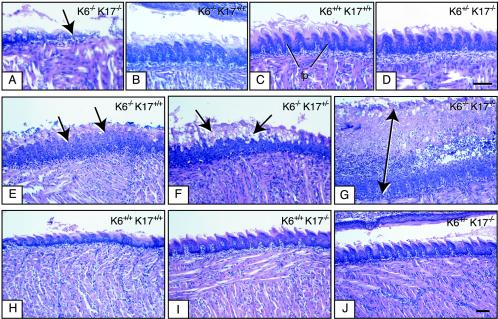
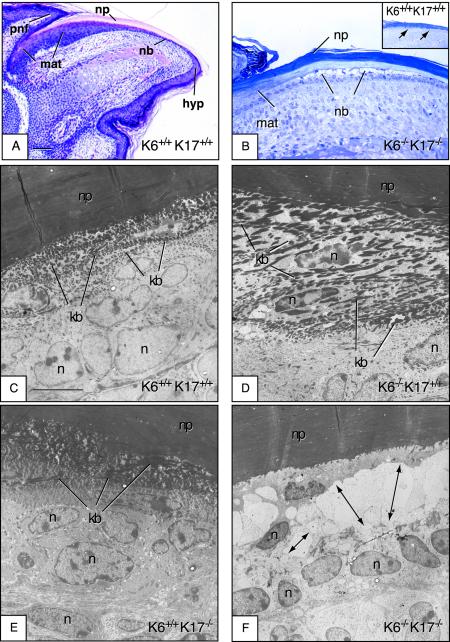
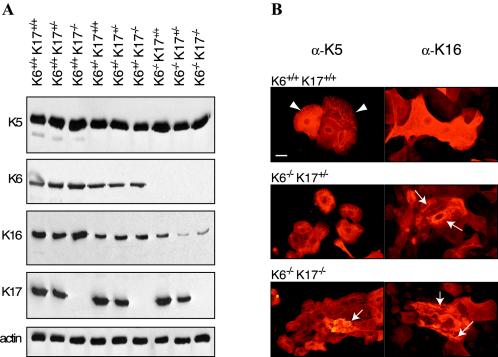
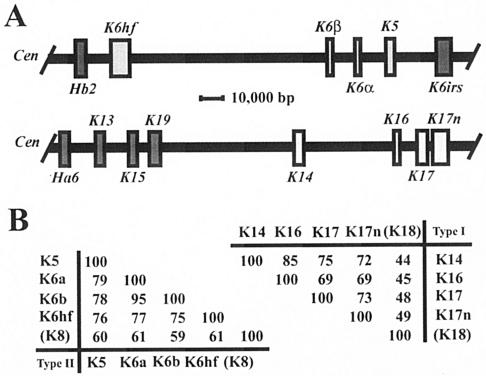
Mutations affecting the coding sequence of intermediate filament (IF) proteins account for >30 disorders, including numerous skin bullous diseases, myopathies, neuropathies, and even progeria. The manipulation of IF genes in mice has been widely successful for modeling key features of such clinically distinct disorders. A notable exception is pachyonychia congenita (PC), a disorder in which the nail and other epithelial appendages are profoundly aberrant. Most cases of PC are due to mutations in one of the following keratin-encoding genes: K6, K16, and K17. Yet null alleles obliterating the function of both K6 genes (K6alpha and K6beta) or the K17 gene, as well as the targeted expression of a dominant-negative K6alpha mutant, elicit only a subset of PC-specific epithelial lesions (excluding that of the nail in mice). We show that newborn mice null for K6alpha, K6beta, and K17 exhibit severe lysis restricted to the nail bed epithelium, where all three genes are robustly expressed, providing strong evidence that this region of the nail unit is initially targeted in PC. Our findings point to significant redundancy among the multiple keratins expressed in hair and nail, which can be related to the common ancestry, clustered organization, and sequence relatedness of specific keratin genes.
Figures
References
-
- Arin, M. J., and D. R. Roop. 2001. Disease model: heritable skin blistering. Trends Mol. Med. 7:422-424. - PubMed
-
- Bernot, K., K. McGowan, and P. A. Coulombe. 2002. Keratin 16 expression defines a subset of epithelial cells during skin morphogenesis and the hair cycle. J. Investig. Dermatol. 119:1137-1149. - PubMed
-
- Blumenberg, M. 1988. Concerted gene duplications in the two keratin gene families. J. Mol. Evol. 27:203-211. - PubMed
-
- Coulombe, P. A., and K. M. Bernot. 2004. Keratins and the skin, p. 497-504. In M. D. Lane, and W. Lennarz, (ed.), Encyclopedia of biological chemistry. Elsevier, Oxford, United Kingdom.
-
- Coulombe, P. A., L. Ma, S. Yamada, and M. Wawersik. 2001. Intermediate filaments at a glance. J. Cell Sci. 114:4345-4347. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous