

A novel intronic cis element, ISE/ISS-3, regulates rat fibroblast growth factor receptor 2 splicing through activation of an upstream exon and repression of a downstream exon containing a noncanonical branch point sequence
- PMID: 15601847
- PMCID: PMC538792
- DOI: 10.1128/MCB.25.1.250-263.2005
A novel intronic cis element, ISE/ISS-3, regulates rat fibroblast growth factor receptor 2 splicing through activation of an upstream exon and repression of a downstream exon containing a noncanonical branch point sequence
Abstract
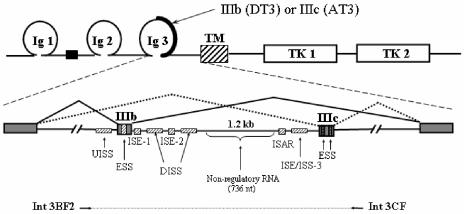
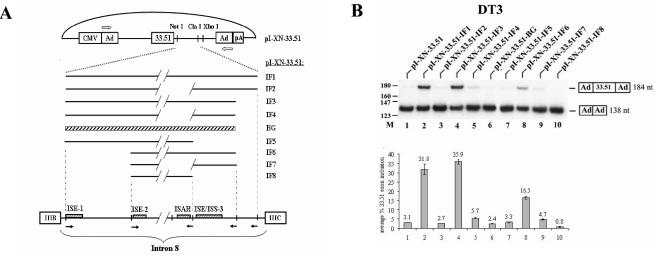
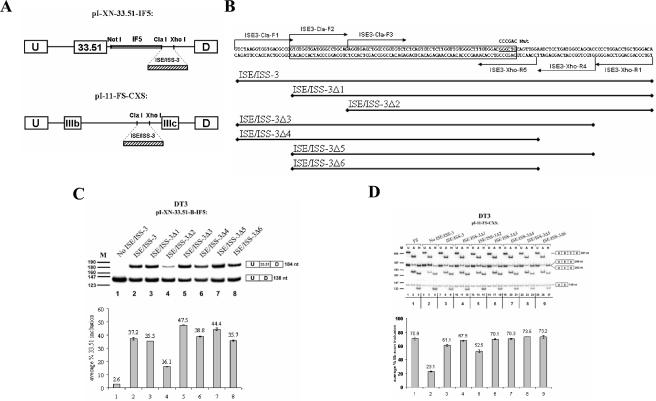
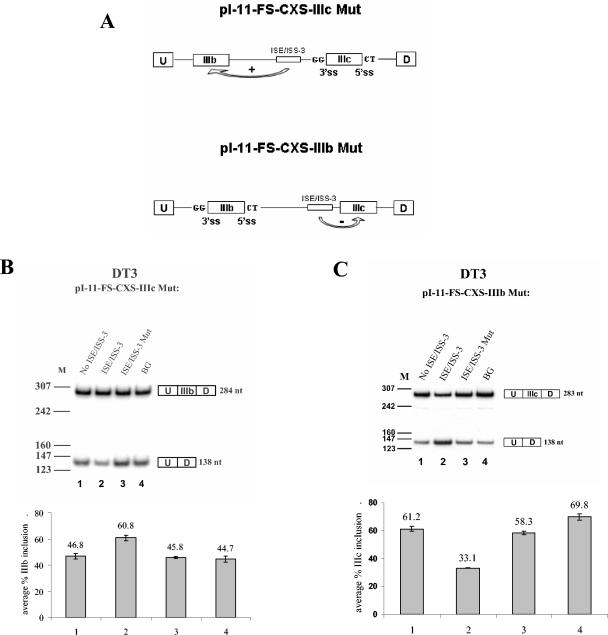
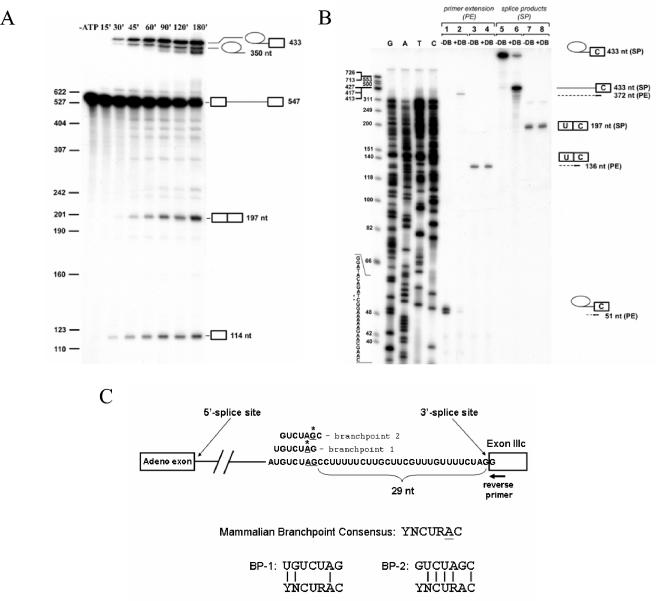
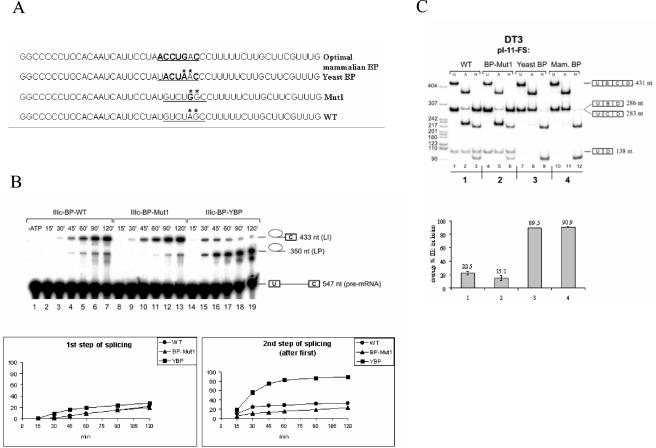
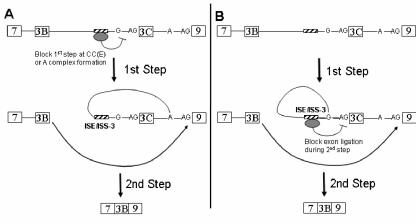
Mutually exclusive splicing of fibroblast growth factor receptor 2 (FGFR2) exons IIIb and IIIc yields two receptor isoforms, FGFR2-IIIb and -IIIc, with distinctly different ligand binding properties. Several RNA cis elements in the intron (intron 8) separating these exons have been described that are required for splicing regulation. Using a heterologous splicing reporter, we have identified a new regulatory element in this intron that confers cell-type-specific inclusion of an unrelated exon that mirrors its ability to promote cell-type-specific inclusion of exon IIIb. This element promoted inclusion of exon IIIb while at the same time silencing exon IIIc inclusion in cells expressing FGFR2-IIIb; hence, we have termed this element ISE/ISS-3 (for "intronic splicing enhancer-intronic splicing silencer 3"). Silencing of exon IIIc splicing by ISE/ISS-3 was shown to require a branch point sequence (BPS) using G as the primary branch nucleotide. Replacing a consensus BPS with A as the primary branch nucleotide resulted in constitutive splicing of exon IIIc. Our results suggest that the branch point sequence constitutes an important component that can contribute to the efficiency of exon definition of alternatively spliced cassette exons. Noncanonical branch points may thus facilitate cell-type-specific silencing of regulated exons by flanking cis elements.
Figures
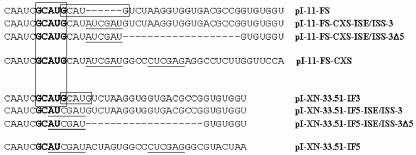
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous