

Conversion of the modulatory actions of dopamine on spinal reflexes from depression to facilitation in D3 receptor knock-out mice
- PMID: 15601940
- PMCID: PMC2731231
- DOI: 10.1523/JNEUROSCI.3698-04.2004
Conversion of the modulatory actions of dopamine on spinal reflexes from depression to facilitation in D3 receptor knock-out mice
Abstract
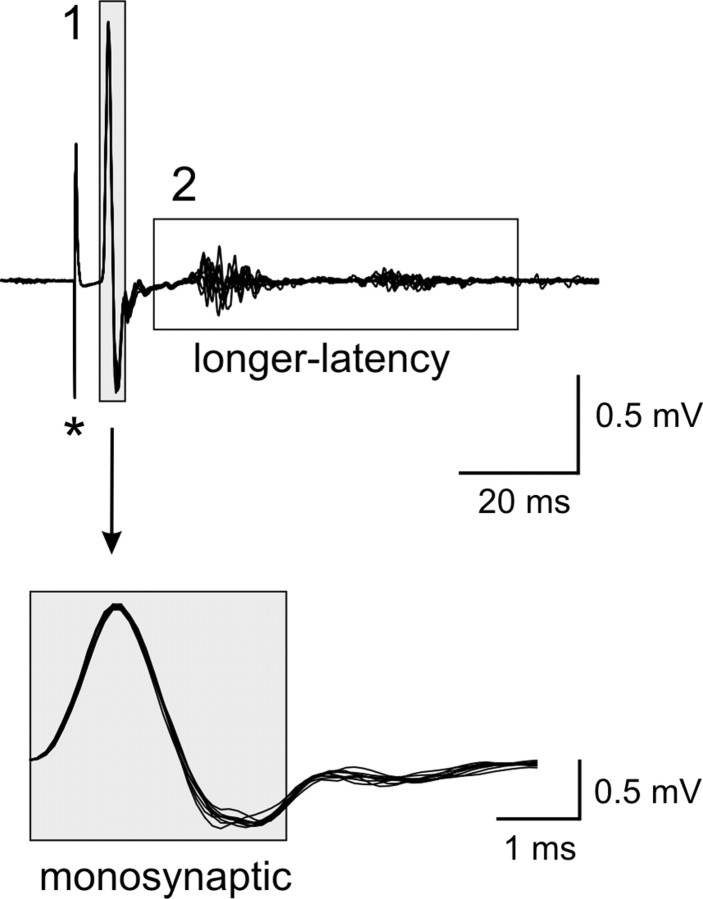
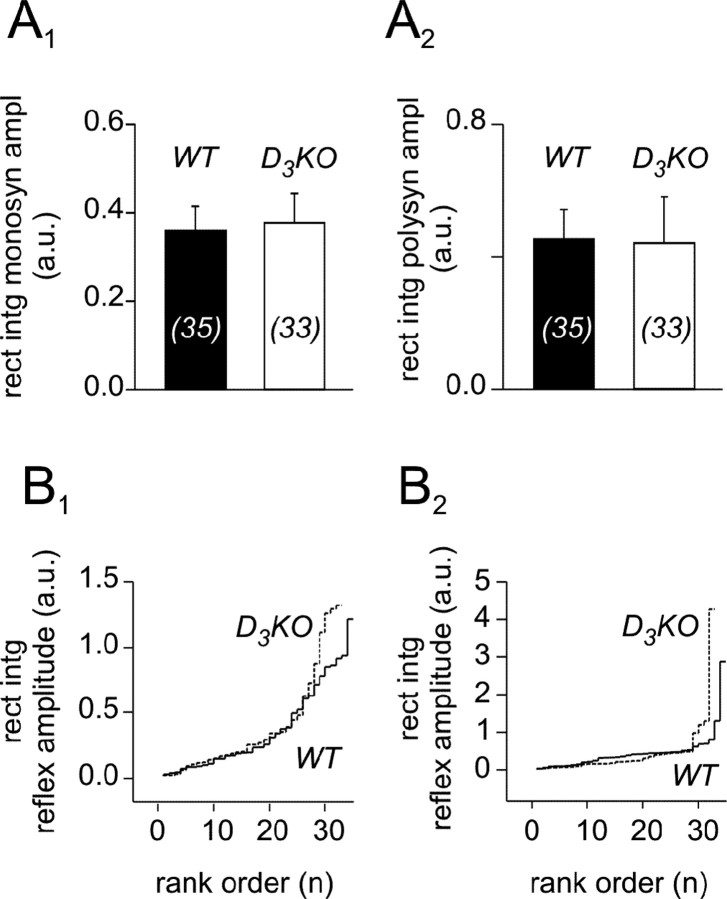
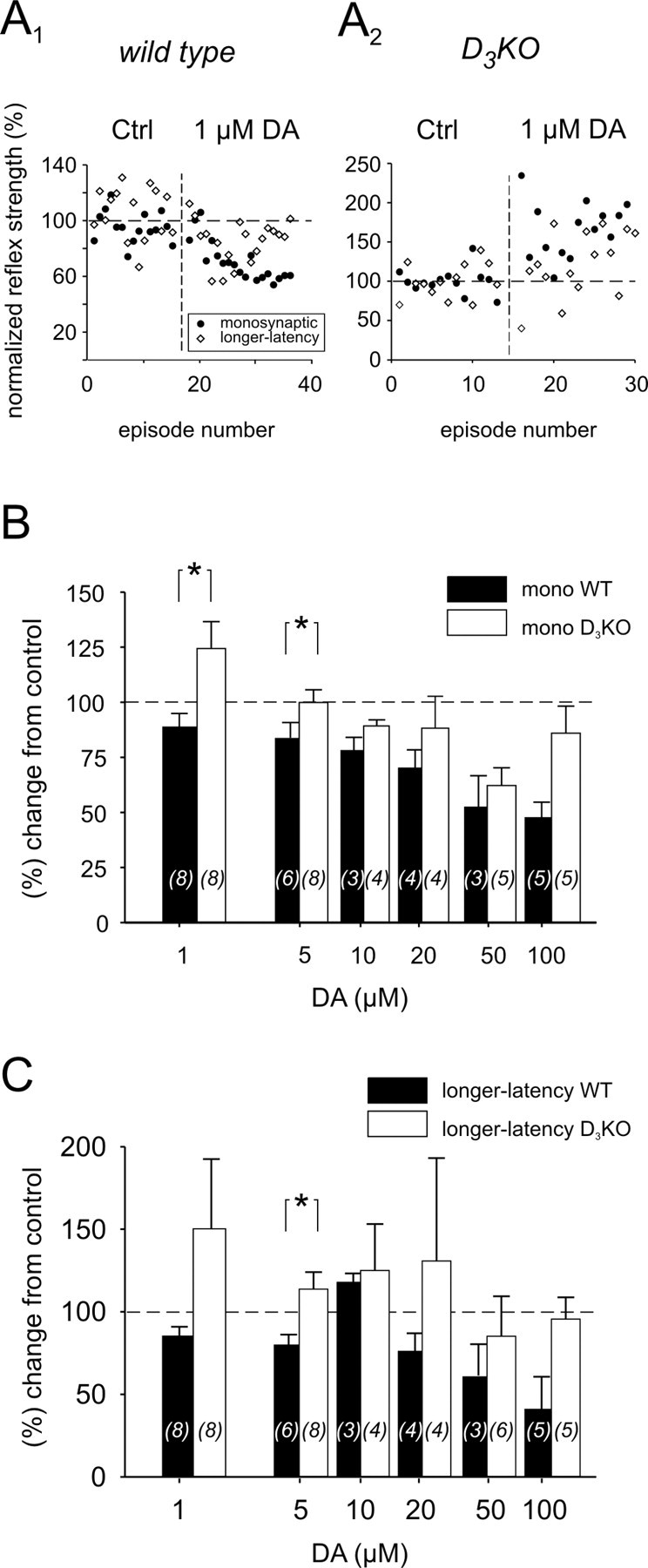
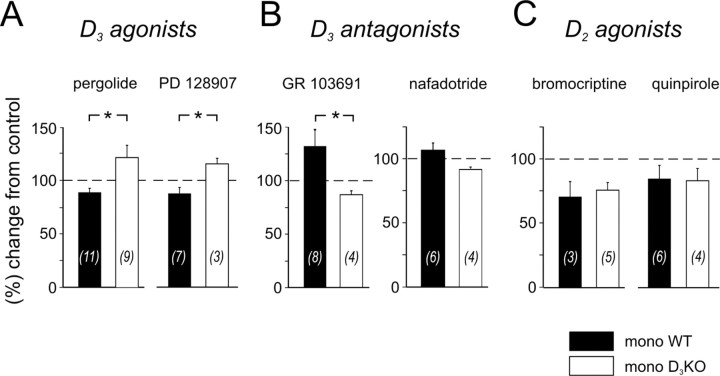
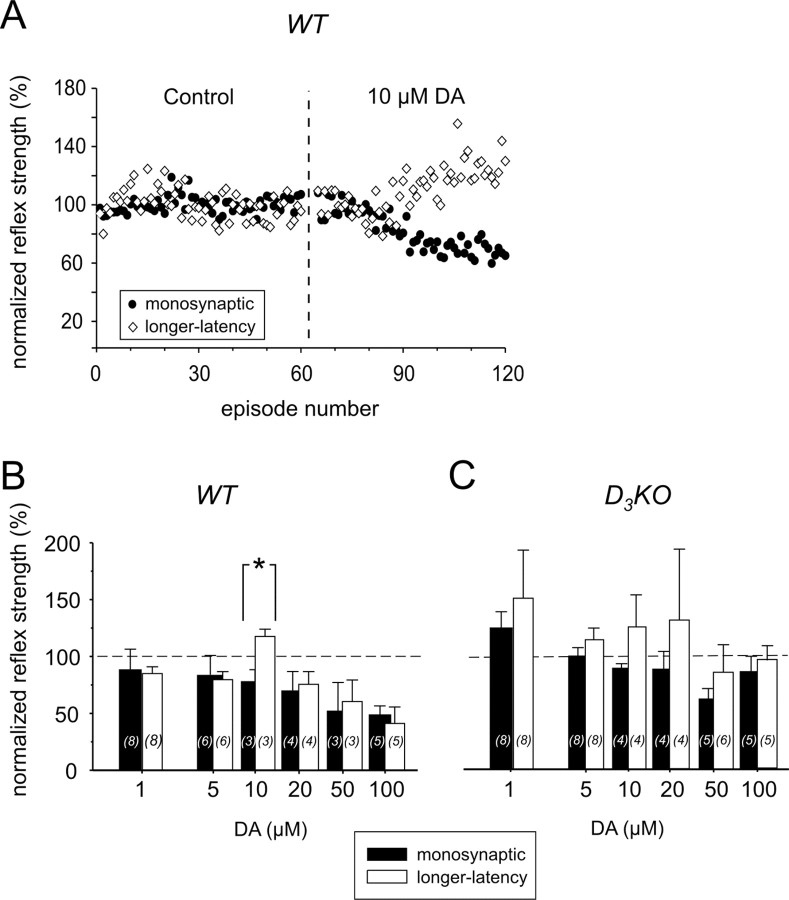
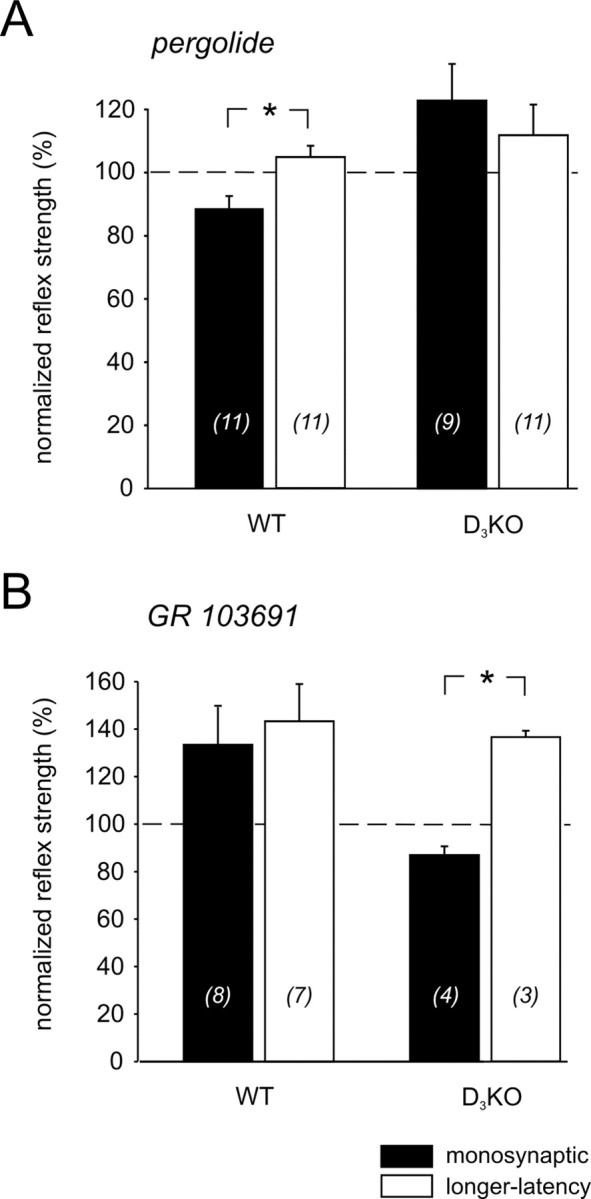
Descending monoaminergic systems modulate spinal cord function, yet spinal dopaminergic actions are poorly understood. Using the in vitro lumbar cord, we studied the effects of dopamine and D2-like receptor ligands on spinal reflexes in wild-type (WT) and D3-receptor knock-out mice (D3KO). Low dopamine levels (1 microM) decreased the monosynaptic "stretch" reflex (MSR) amplitude in WT animals and increased it in D3KO animals. Higher dopamine concentrations (10-100 microM) decreased MSR amplitudes in both groups, but always more strongly in WT. Like low dopamine, the D3 receptor agonists pergolide and PD 128907 reduced MSR amplitude in WT but not D3KO mice. Conversely, D3 receptor antagonists (GR 103691 and nafadotride) increased the MSR in WT but not in D3KO mice. In comparison, D2-preferring agonists bromocriptine and quinpirole depressed the MSR in both groups. Low dopamine (1-5 microM) also depressed longer-latency (presumably polysynaptic) reflexes in WT but facilitated responses in D3KO mice. Additionally, in some experiments (e.g., during 10 microM dopamine or pergolide in WT), polysynaptic reflexes were facilitated in parallel to MSR depression, demonstrating differential modulatory control of these reflex circuits. Thus, low dopamine activates D3 receptors to limit reflex excitability. Moreover, in D3 ligand-insensitive mice, excitatory actions are unmasked, functionally converting the modulatory action of dopamine from depression to facilitation. Restless legs syndrome (RLS) is a CNS disorder involving abnormal limb sensations. Because RLS symptoms peak at night when dopamine levels are lowest, are relieved by D3 agonists, and likely involve increased reflex excitability, the D3KO mouse putatively explains how impaired D3 activity could contribute to this sleep disorder.
Figures
References
-
- Abramets II, Samoilovich IM (1991) Analysis of two types of dopaminergic responses of neurons of the spinal ganglia of rats. Neurosci Behav Physiol 21: 435-440. - PubMed
-
- Ali NJ, Davies RJ, Fleetham JA, Stradling JR (1991) Periodic movements of the legs during sleep associated with rises in systemic blood pressure. Sleep 14: 163-165. - PubMed
-
- Allen RP, Earley CJ (2001) Restless legs syndrome: a review of clinical and pathophysiologic features. J Clin Neurophysiol 18: 128-147. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases