

Interaction between the corticotropin-releasing factor system and hypocretins (orexins): a novel circuit mediating stress response
- PMID: 15601950
- PMCID: PMC6730356
- DOI: 10.1523/JNEUROSCI.3459-04.2004
Interaction between the corticotropin-releasing factor system and hypocretins (orexins): a novel circuit mediating stress response
Abstract
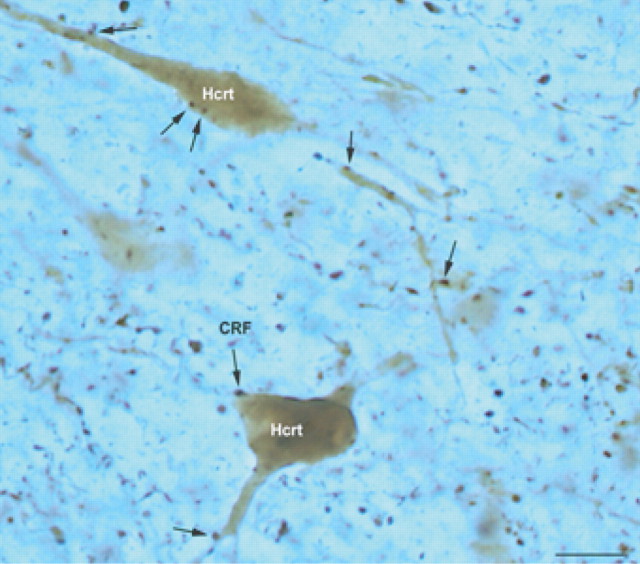
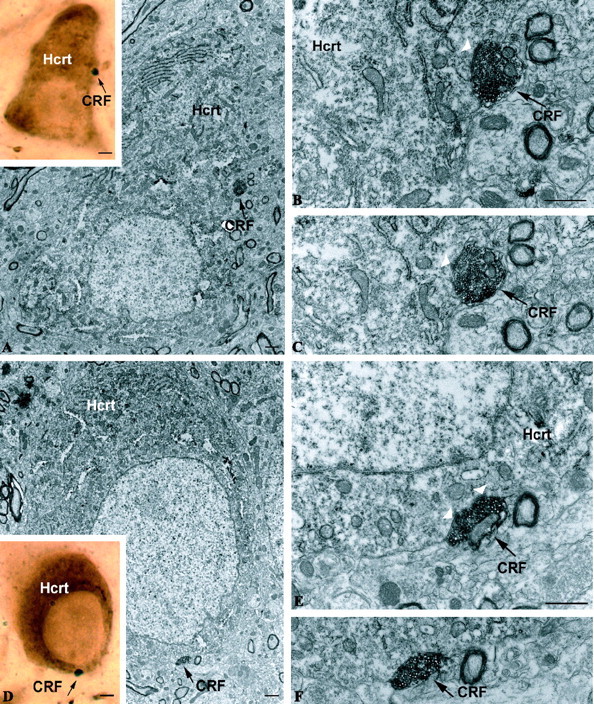
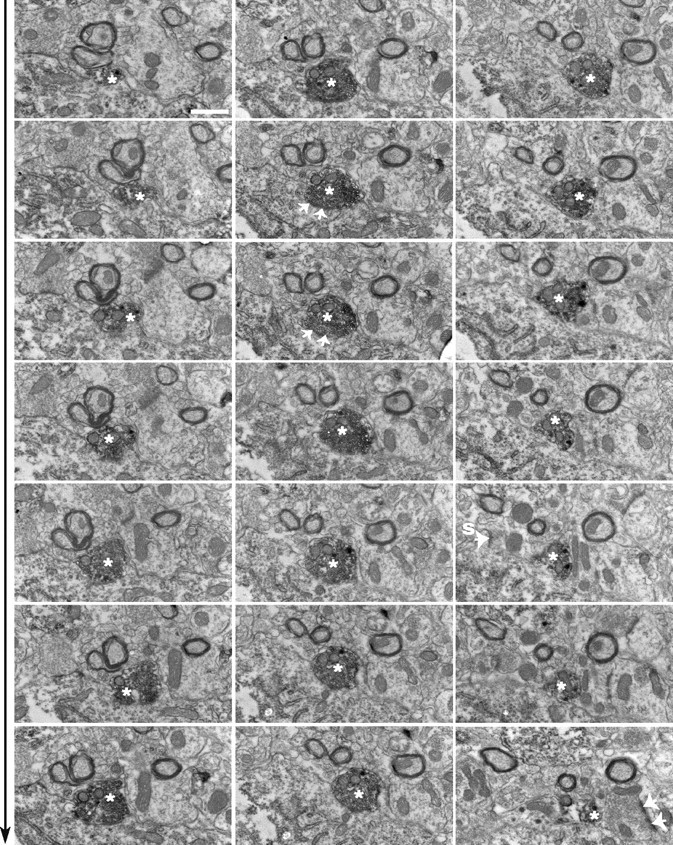
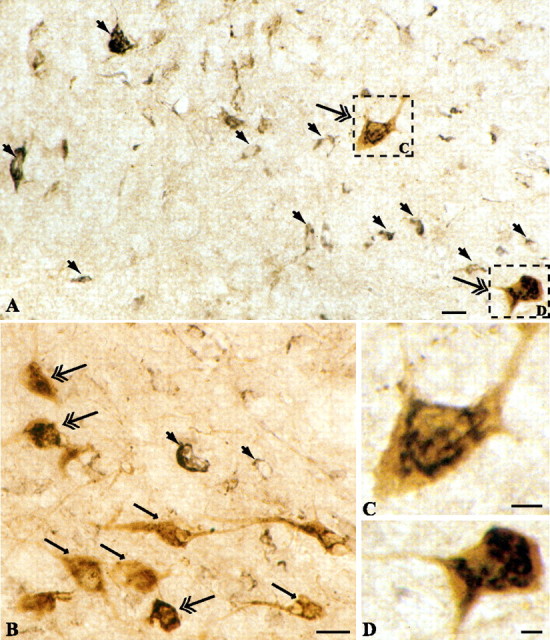
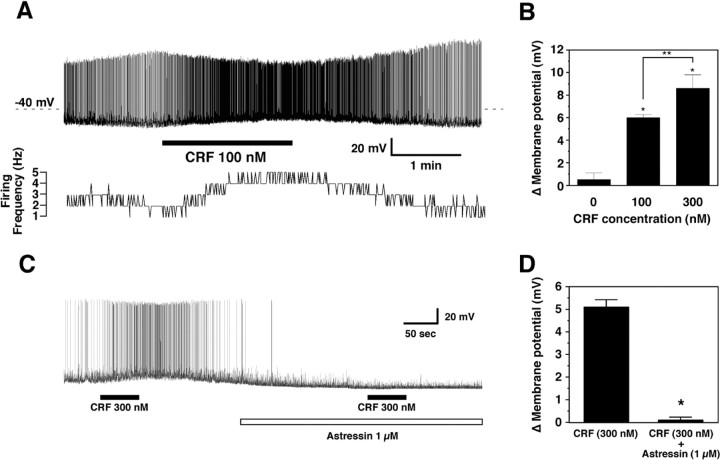
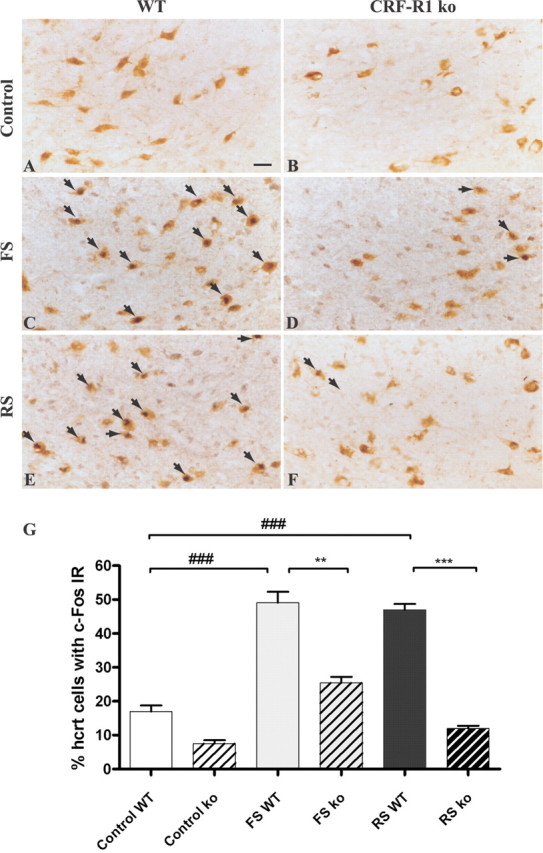
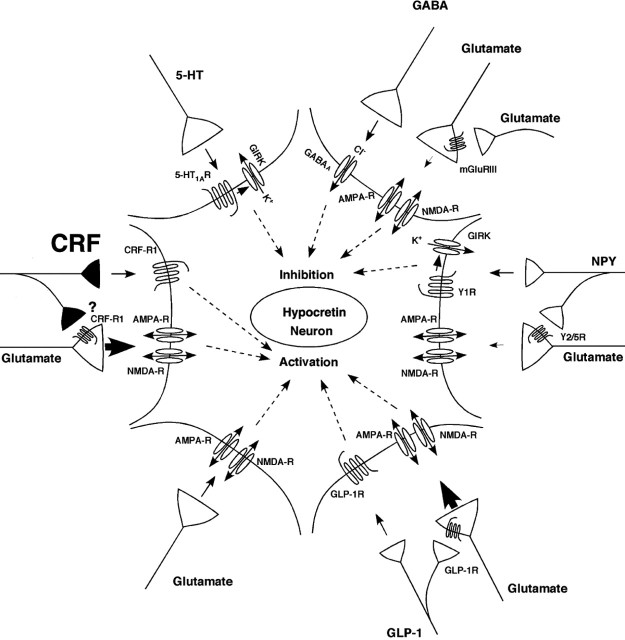
The hypothalamic neuropeptides hypocretins (orexins) play a crucial role in the stability of arousal and alertness. We tested whether the hypocretinergic system is a critical component of the stress response activated by the corticotropin-releasing factor (CRF). Our results show that CRF-immunoreactive terminals make direct contact with hypocretin-expressing neurons in the lateral hypothalamus and that numerous hypocretinergic neurons express the CRF-R1/2 receptors. We also demonstrate that application of CRF to hypothalamic slices containing identified hypocretin neurons depolarizes membrane potential and increases firing rate in a subpopulation of hypocretinergic cells. CRF-induced depolarization was tetrodotoxin insensitive and was blocked by the peptidergic CRF-R1 antagonist astressin. Moreover, activation of hypocretinergic neurons in response to acute stress was severely impaired in CRF-R1 knock-out mice. Together, our data provide evidence of a direct neuroanatomical and physiological input from CRF peptidergic system onto hypocretin neurons. We propose that, after stressor stimuli, CRF stimulates the release of hypocretins and that this circuit contributes to activation and maintenance of arousal associated with the stress response.
Figures

References
-
- Blanco M, Lopez M, GarcIa-Caballero T, Gallego R, Vazquez-Boquete A, Morel G, SenarIs R, Casanueva F, Dieguez C, Beiras A (2001) Cellular localization of orexin receptors in human pituitary. J Clin Endocrinol Metab 86: 1616-1619. - PubMed
-
- Blanco M, Gallego R, Garcia-Caballero T, Dieguez C, Beiras A (2003) Cellular localization of orexins in human anterior pituitary. Histochem Cell Biol 120: 259-264. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases