

The role of chemokines and extracellular matrix components in the migration of T lymphocytes into three-dimensional substrata
- PMID: 15606795
- PMCID: PMC1782061
- DOI: 10.1111/j.1365-2567.2004.02005.x
The role of chemokines and extracellular matrix components in the migration of T lymphocytes into three-dimensional substrata
Abstract
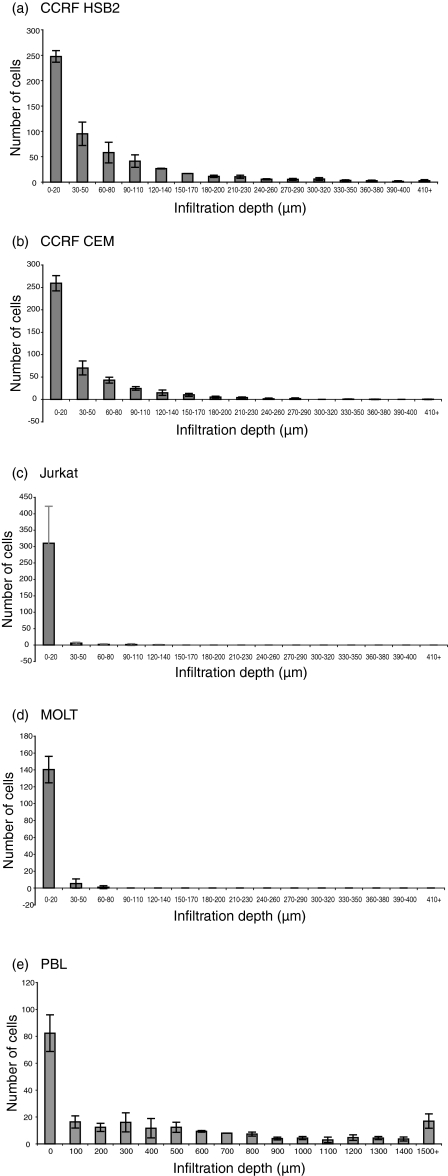
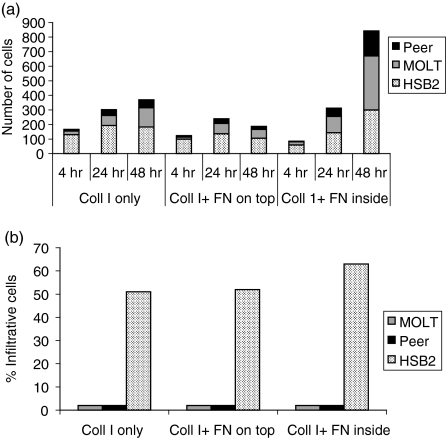
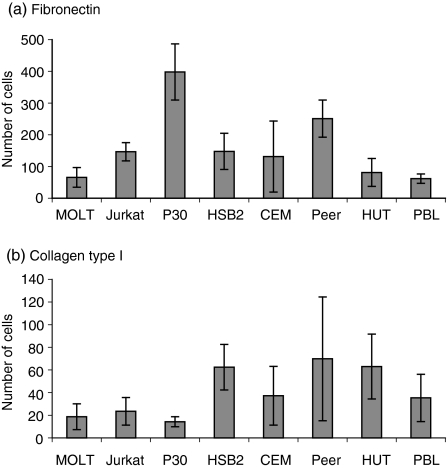
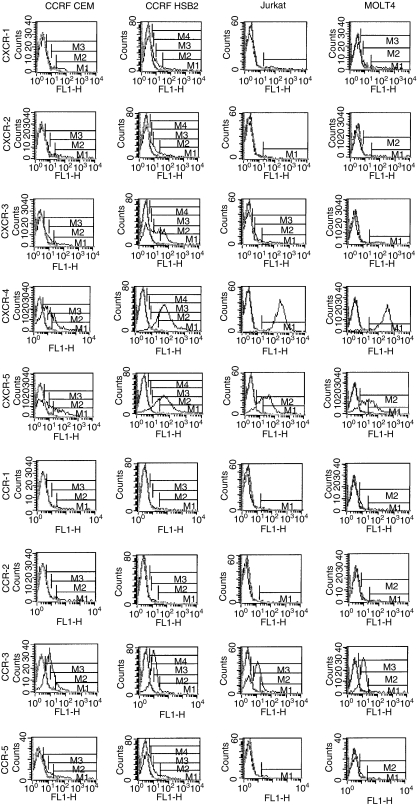
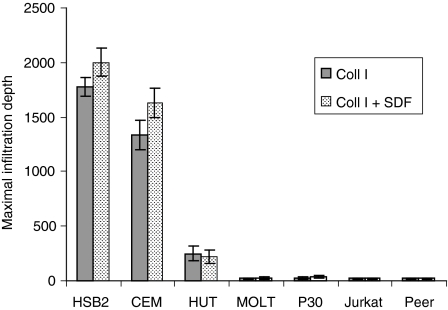
The role of chemokines and their interactions with extracellular matrix components (ECM) or the capacity of T cells to migrate into and accumulate within three-dimensional (3D) collagen type 1 substrata was studied. We examined the influence of chemokines and fibronectin on the infiltration properties of non-infiltrative (do not migrate into 3D substrata) and spontaneously infiltrative (migrate into 3D substrata) T-cell lines. Infiltrative and non-infiltrative T-acute lymphocytic leukaemic cell lines exhibited no consistent differences with respect to the expression of various chemokine receptors or beta(1)-integrins. Chemokines presented inside the collagen increased the depth of migration of infiltrative T-cell lines, but did not render non-infiltrative T-cell lines infiltrative, although they augmented the attachment of non-infiltrative T-cell lines to the upper surface of the collagen. The presence of fibronectin inside the collagen did not render non-infiltrative T-cell lines infiltrative, but markedly augmented the migration of 'infiltrative' T-cell lines into collagen. Both infiltrative and non-infiltrative T-cell lines showed migratory responses to chemokines in Boyden assays (migration detected on 2D substrata). These results indicate that the process of T-cell infiltration/migration into 3D substrata depends on a tissue penetration mechanism distinguishable from migration on 2D substrata and that the basic capacity of T cells to infiltrate is independent of chemokines and ECM components applied as attractants.
Figures

References
-
- Hauzenberger D, Klominek J, Bergström SE, Sundqvist KG. T lymphocyte migration: the influence of interactions via adhesion molecules, the T cell receptor and cytokines. Crit Rev Immunol. 1995;15:285–316. - PubMed
-
- Lawrence MB, Springer TA. Leukocytes roll on a selectin at physiologic flow rates: distinction from and prerequisite for adhesion through integrins. Cell. 1991;65:859–73. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
