

Analysis of 2'-phosphotransferase (Tpt1p) from Saccharomyces cerevisiae: evidence for a conserved two-step reaction mechanism
- PMID: 15611300
- PMCID: PMC1370695
- DOI: 10.1261/rna.7194605
Analysis of 2'-phosphotransferase (Tpt1p) from Saccharomyces cerevisiae: evidence for a conserved two-step reaction mechanism
Abstract
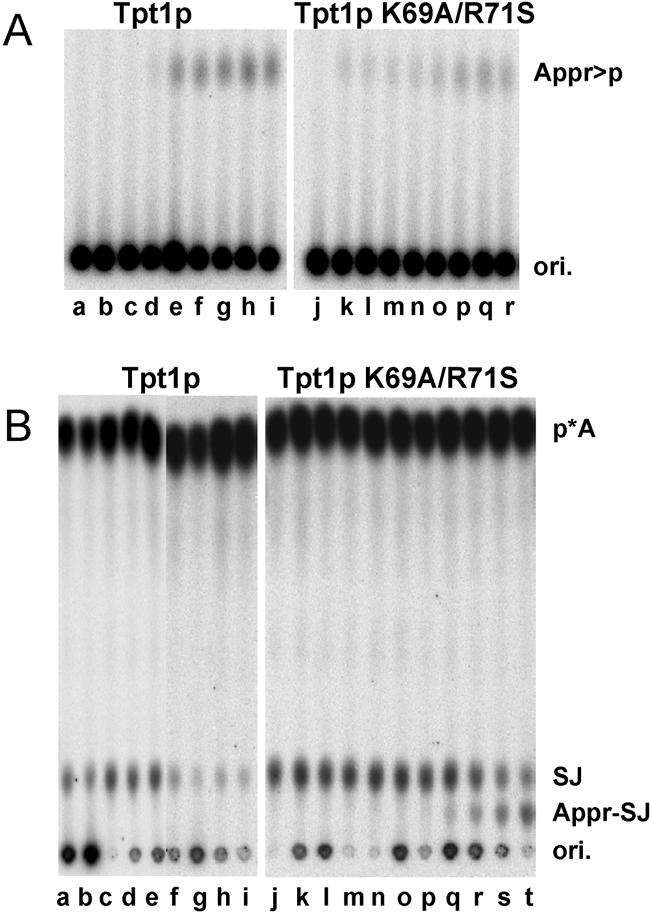
Tpt1p is an essential protein responsible for the 2'-phosphotransferase step of tRNA splicing in Saccharomyces cerevisiae, in which the splice junction 2'-phosphate of ligated tRNA is transferred to NAD to form mature tRNA and ADP-ribose 1''-2'' cyclic phosphate. We showed previously that Tpt1p is a member of a family of functional 2'-phosphotransferases found in eukaryotes, eubacteria, and archaea, that the Escherichia coli protein (KptA) is highly specific for 2'-phosphorylated RNAs despite the lack of obvious natural substrates, and that KptA acts on a trinucleotide substrate through an intermediate in which RNA is ADP-ribosylated at the 2'-phosphate. This mechanism is similar to a proposed mechanism of NAD-dependent histone deacetylases. We present evidence here that this mechanism is conserved in S. cerevisiae, and we identify residues important for the second step of the reaction, during which the intermediate is resolved into products. We examined 21 Tpt1 protein variants mutated in conserved residues or blocks of residues and show that one of them, Tpt1 K69A/R71S protein, accumulates large amounts of intermediate with trinucleotide substrate due to a very slow second step. This intermediate can be trapped on beads when formed with biotin-NAD. We also show that Tpt1 K69A/R71S protein forms an intermediate with the natural ligated tRNA substrate and demonstrate that, as expected, this mutation is lethal in yeast. The high degree of conservation of these residues suggests that the entire Tpt1p family is involved in a similar two-step chemical reaction.
Figures






References
-
- Abelson, J., Trotta, C.R., and Li, H. 1998. tRNA splicing. J. Biol. Chem. 273: 12685–12688. - PubMed
-
- Barnard, E.A. 1969. Ribonucleases. Annu. Rev. BioChem. 38: 677–732. - PubMed
-
- Belford, H.G., Westaway, S.K., Abelson, J., and Greer, C.L. 1993. Multiple nucleotide cofactor use by yeast ligase in tRNA splicing. Evidence for independent ATP- and GTP-binding sites. J. Biol. Chem. 268: 2444–2450. - PubMed
-
- Bell, C.E. and Eisenberg, D. 1996. Crystal structure of diphtheria toxin bound to nicotinamide adenine dinucleotide. Biochemistry 35: 1137–1149. - PubMed
-
- Bradford, M.M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. BioChem. 72: 248–254. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases