

In vitro and in vivo mutational analysis of the 3'-terminal regions of hepatitis e virus genomes and replicons
- PMID: 15613330
- PMCID: PMC538530
- DOI: 10.1128/JVI.79.2.1017-1026.2005
In vitro and in vivo mutational analysis of the 3'-terminal regions of hepatitis e virus genomes and replicons
Abstract
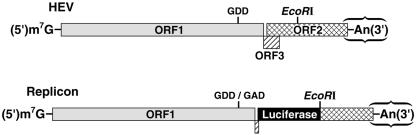
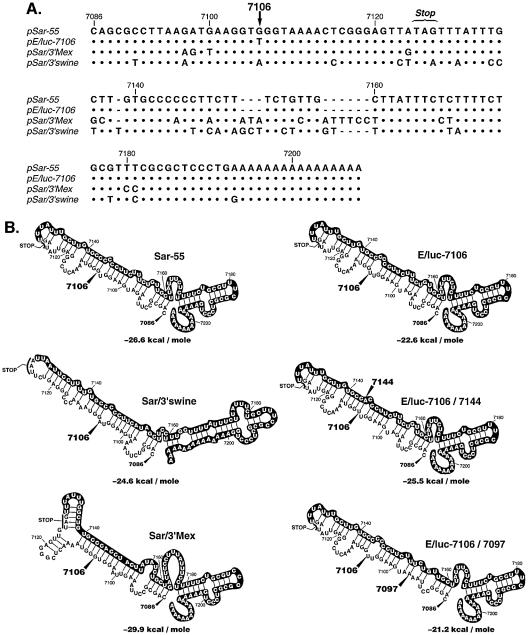
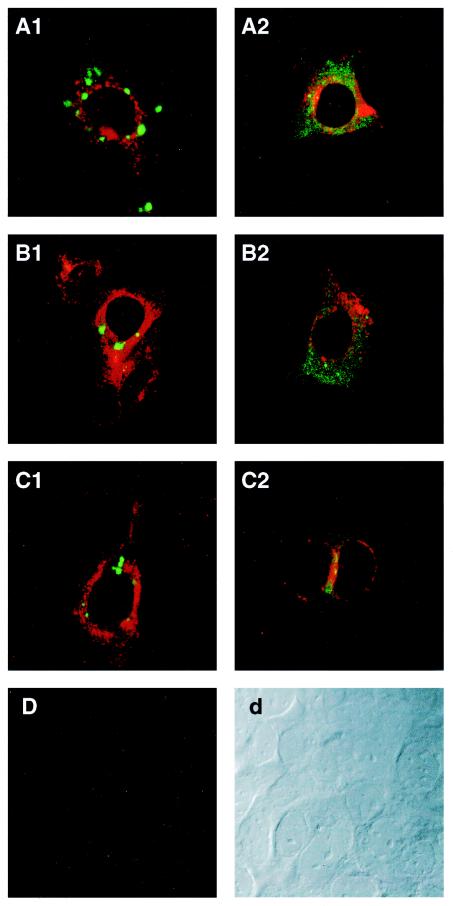
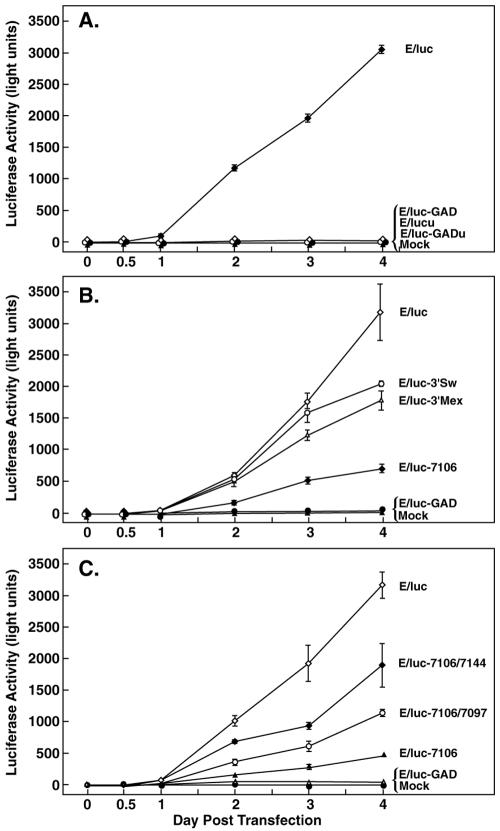
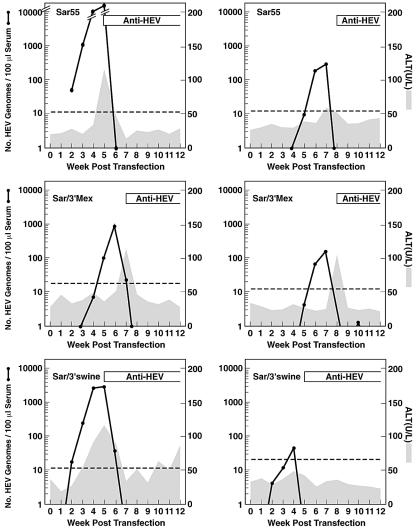
Hepatitis E virus (HEV) replication is not well understood, mainly because the virus does not infect cultured cells efficiently. However, Huh-7 cells transfected with full-length genomes produce open reading frame 2 protein, indicative of genome replication (6). To investigate the role of 3'-terminal sequences in RNA replication, we constructed chimeric full-length genomes with divergent 3'-terminal sequences of genotypes 2 and 3 replacing that of genotype 1 and transfected them into Huh-7 cells. The production of viral proteins by these full-length chimeras was indistinguishable from that of the wild type, suggesting that replication was not impaired. In order to better quantify HEV replication in cell culture, we constructed an HEV replicon with a reporter (luciferase). Luciferase production was cap dependent and RNA-dependent RNA polymerase dependent and increased following transfection of Huh-7 cells. Replicons harboring the 3'-terminal intergenotypic chimera sequences were also assayed for luciferase production. In spite of the large sequence differences among the 3' termini of the viruses, replication of the chimeric replicons was surprisingly similar to that of the parental replicon. However, a single unique nucleotide change within a predicted stem structure at the 3' terminus substantially reduced the efficiency of replication: RNA replication was partially restored by a covariant mutation. Similar patterns of replication were obtained when full-length genomes were inoculated into rhesus macaques, suggesting that the in vitro system could be used to predict the effect of 3'-terminal mutations in vivo. Incorporation of the 3'-terminal sequences of the swine strain of HEV into the genotype 1 human strain did not enable the human strain to infect swine.
Figures
References
-
- Agrawal, S., D. Gupta, and S. K. Panda. 2001. The 3′ end of hepatitis E virus (HEV) genome binds specifically to the viral RNA-dependent RNA polymerase (RdRp). Virology 282:87-101. - PubMed
-
- Arankalle, V. A., L. P. Chobe, A. M. Walimbe, P. N. Yergolkar, and G. P. Jacob. 2003. Swine HEV infection in south India and phylogenetic analysis (1985-1999). J. Med. Virol. 69:391-396. - PubMed
-
- Bradley, D. W., K. Krawczynski, E. H. Cook, Jr., K. A. McCaustland, C. D. Humphrey, J. E. Spelbring, H. Myint, and J. E. Maynard. 1987. Enterically transmitted non-A, non-B hepatitis: serial passage of disease in cynomolgus macaques and tamarins and recovery of disease-associated 27- to 34-nm viruslike particles. Proc. Natl. Acad. Sci. USA 84:6277-6281. - PMC - PubMed
-
- Dollenmaier, G., and M. Weitz. 2003. Interaction of glyceraldehyde-3-phosphate dehydrogenase with secondary and tertiary RNA structural elements of the hepatitis A virus 3′ translated and non-translated regions. J. Gen. Virol. 84:403-414. - PubMed
-
- Emerson, S. U., D. Anderson, A. Arankalle, X.-J. Meng, M. Purdy, G. G. Schlauder, and S. A. Tsarev. 2004. Hepevirus, p. 851-855. In C. M. Fauquet, M. A. Mayo, J. Maniloff, U. Desselberger, and L. A. Ball (ed.), Virus taxonomy. VIIIth report of the ICTV. Elsevier/Academic Press, London, United Kingdom.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
