

Antigenicity and immunogenicity of a synthetic human immunodeficiency virus type 1 group m consensus envelope glycoprotein
- PMID: 15613343
- PMCID: PMC538535
- DOI: 10.1128/JVI.79.2.1154-1163.2005
Antigenicity and immunogenicity of a synthetic human immunodeficiency virus type 1 group m consensus envelope glycoprotein
Abstract
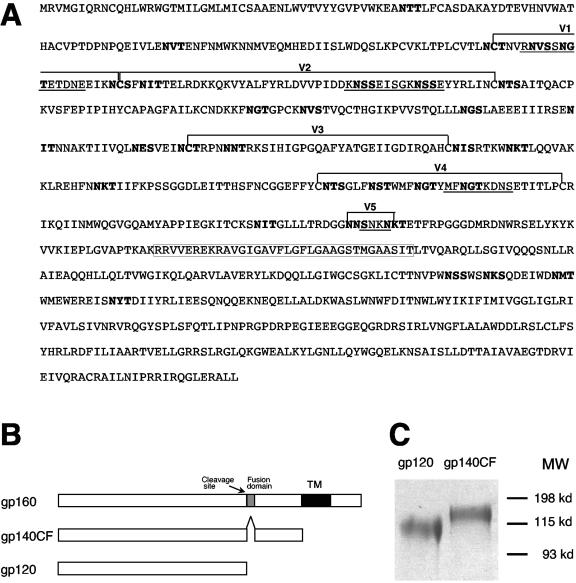
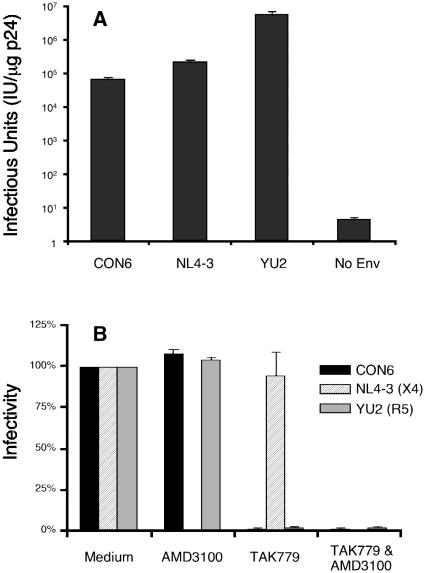
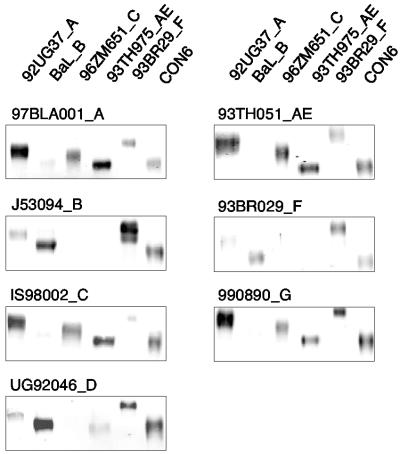
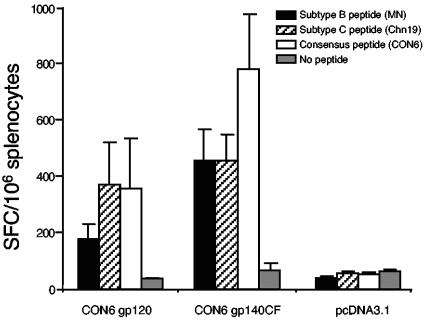
Genetic variation of human immunodeficiency virus (HIV-1) represents a major obstacle for AIDS vaccine development. To decrease the genetic distances between candidate immunogens and field virus strains, we have designed and synthesized an artificial group M consensus env gene (CON6 gene) to be equidistant from contemporary HIV-1 subtypes and recombinants. This novel envelope gene expresses a glycoprotein that binds soluble CD4, utilizes CCR5 but not CXCR4 as a coreceptor, and mediates HIV-1 entry. Key linear, conformational, and glycan-dependent monoclonal antibody epitopes are preserved in CON6, and the glycoprotein is recognized equally well by sera from individuals infected with different HIV-1 subtypes. When used as a DNA vaccine followed by a recombinant vaccinia virus boost in BALB/c mice, CON6 env gp120 and gp140CF elicited gamma interferon-producing T-cell responses that recognized epitopes within overlapping peptide pools from three HIV-1 Env proteins, CON6, MN (subtype B), and Chn19 (subtype C). Sera from guinea pigs immunized with recombinant CON6 Env gp120 and gp140CF glycoproteins weakly neutralized selected HIV-1 primary isolates. Thus, the computer-generated "consensus" env genes are capable of expressing envelope glycoproteins that retain the structural, functional, and immunogenic properties of wild-type HIV-1 envelopes.
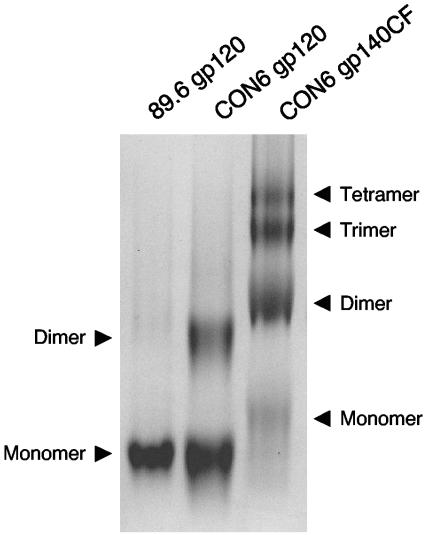
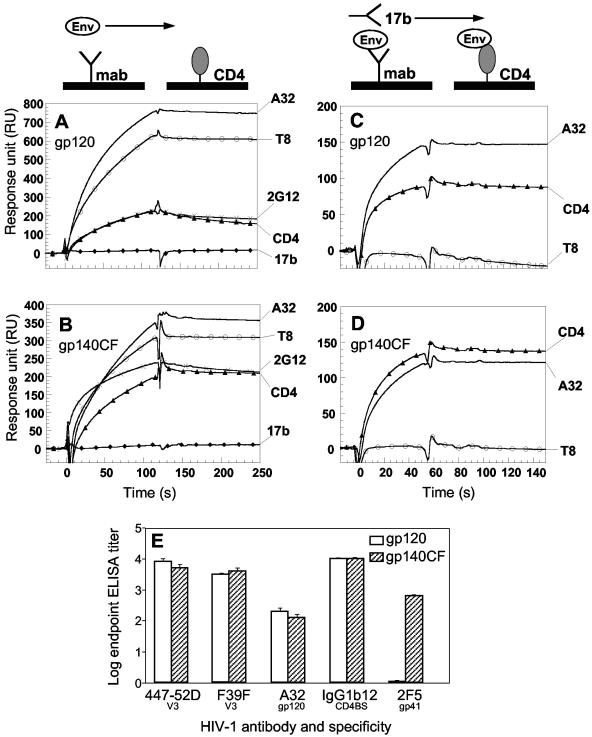
Figures
References
-
- Brandt, S. M., R. Mariani, A. U. Holland, T. J. Hope, and N. R. Landau. 2002. Association of chemokine-mediated block to HIV entry with coreceptor internalization. J. Biol. Chem. 277:17291-17299. - PubMed
-
- Bures, R., A. Gaitan, T. Zhu, C. Graziosi, K. M. McGrath, J. Tartaglia, P. Caudrelier, R. El Habib, M. Klein, A. Lazzarin, D. M. Stablein, M. Deers, L. Corey, M. L. Greenberg, D. H. Schwartz, and D. C. Montefiori. 2000. Immunization with recombinant canarypox vectors expressing membrane-anchored glycoprotein 120 followed by glycoprotein 160 boosting fails to generate antibodies that neutralize R5 primary isolates of human immunodeficiency virus type 1. AIDS Res. Hum. Retroviruses 16:2019-2035. - PubMed
-
- Carr, J. K., B. T. Foley, T. Leitner, M. O. Salminen, B. T. Korber, and F. McCutchan. 1998. Reference sequences representing the principal genetic diversity of HIV-1 in the pandemic, p. III-10-III-19. In B. Korber, C. Kuiken, B. Foley, B. H. Hahn, F. McCutchan, J. W. Mellors, and J. Sodrosky (ed.), Human retroviruses and AIDS 1998: a compilation and analysis of nucleic acid and amino acid sequence. Theoretical Biology and Biophysics Group, Los Alamos National Laboratory, Los Alamos, N.Mex.
Publication types
MeSH terms
Substances
Grants and funding
- T32 AI007392/AI/NIAID NIH HHS/United States
- AI55386/AI/NIAID NIH HHS/United States
- P0-1 AI52816/AI/NIAID NIH HHS/United States
- N01 AI085338/AI/NIAID NIH HHS/United States
- R21 AI055386/AI/NIAID NIH HHS/United States
- P0-1 AI51445/AI/NIAID NIH HHS/United States
- U01 AI035351/AI/NIAID NIH HHS/United States
- AI54497/AI/NIAID NIH HHS/United States
- 5T32 AI07392/AI/NIAID NIH HHS/United States
- AI35351/AI/NIAID NIH HHS/United States
- P01 AI035351/AI/NIAID NIH HHS/United States
- AI85338/AI/NIAID NIH HHS/United States
- P30 AI051445/AI/NIAID NIH HHS/United States
- P01 AI052816/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
