

Generation of active pools of abscisic acid revealed by in vivo imaging of water-stressed Arabidopsis
- PMID: 15618419
- PMCID: PMC548852
- DOI: 10.1104/pp.104.053082
Generation of active pools of abscisic acid revealed by in vivo imaging of water-stressed Arabidopsis
Abstract
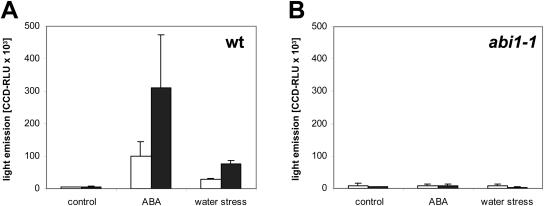
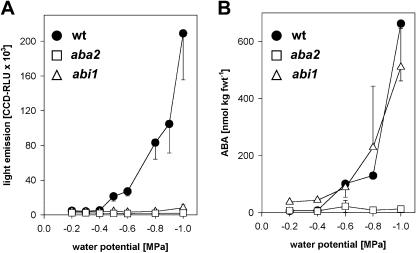
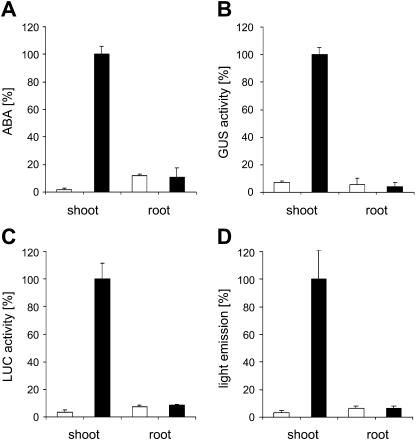
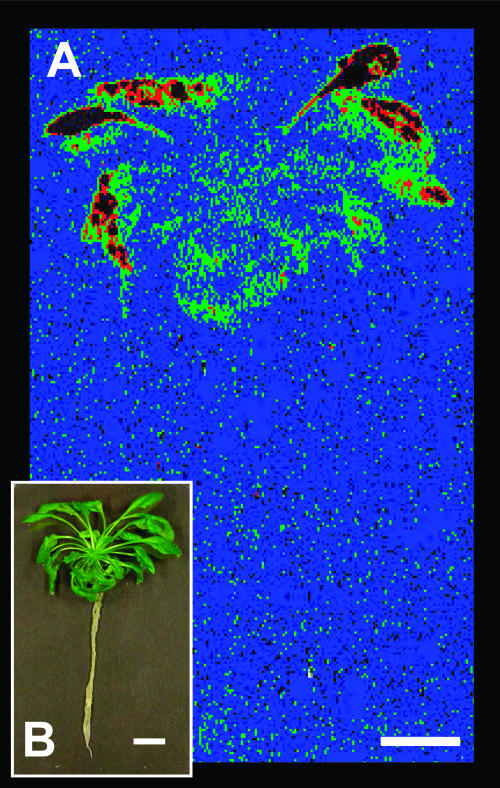
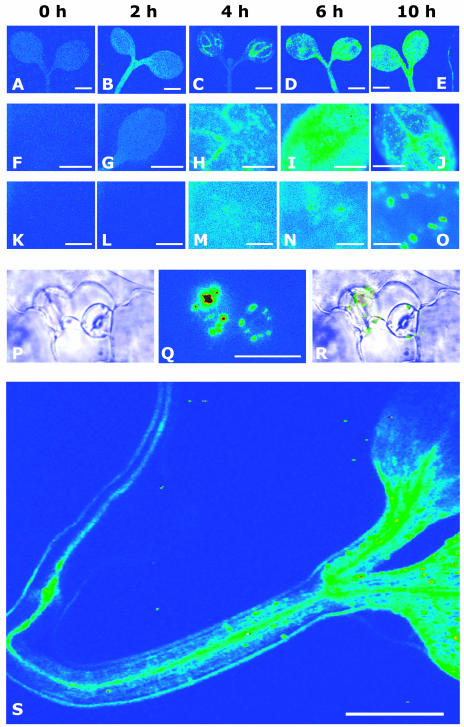
A noninvasive, cell-autonomous reporter system was developed to monitor the generation and distribution of physiologically active pools of abscisic acid (ABA). ABA response (abi1-1) and biosynthesis (aba2-1) mutants of Arabidopsis (Arabidopsis thaliana) were used to validate the system in the presence and absence of water stress. In the absence of water stress, low levels of ABA-dependent reporter activation were observed in the columella cells and quiescent center of the root as well as in the vascular tissues and stomata of cotyledons, suggesting a nonstress-related role for ABA in these cell types. Exposure of seedlings to exogenous ABA resulted in a uniform pattern of reporter expression. In marked contrast, reporter expression in response to drought stress was predominantly confined to the vasculature and stomata. Surprisingly, water stress applied to the root system resulted in the generation of ABA pools in the shoot but not in the root. The analysis of the response dynamics revealed a spread of physiologically active ABA from the vascular tissue into the areoles of the cotyledons. Later, ABA preferentially activated gene expression in guard cells. The primary sites of ABA action identified by in planta imaging corresponded to the sites of ABA biosynthesis, i.e. guard cells and cells associated with vascular veins. Hence, water stress recognized by the root system predominantly results in shoot-localized ABA action that culminates in a focused response in guard cells.
Figures



References
-
- Benkova E, Michniewicz M, Sauer M, Teichmann T, Seifertova D, Jurgens G, Friml J (2003) Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 115: 591–602 - PubMed
-
- Breyne P, De Loose M, Dedonder A, Van Montagu M, Depicker A (1993) Quantitative analysis of β-glucuronidase activities using a computer-directed microtiter plate reader. Plant Mol Biol Rep 11: 21–31
-
- Casimiro I, Beeckman T, Graham N, Bhalerao R, Zhang H, Casero P, Sandberg G, Bennett MJ (2003) Dissecting Arabidopsis lateral root development. Trends Plant Sci 8: 165–171 - PubMed
-
- Chinnusamy V, Stevenson B, Lee BH, Zhu JK (2002) Screening for gene regulation mutants by bioluminescence imaging. Sci STKE 140: PL10 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
