

Phosphate forms an unusual tripodal complex with the Fe-Mn center of sweet potato purple acid phosphatase
- PMID: 15625111
- PMCID: PMC544300
- DOI: 10.1073/pnas.0407239102
Phosphate forms an unusual tripodal complex with the Fe-Mn center of sweet potato purple acid phosphatase
Abstract
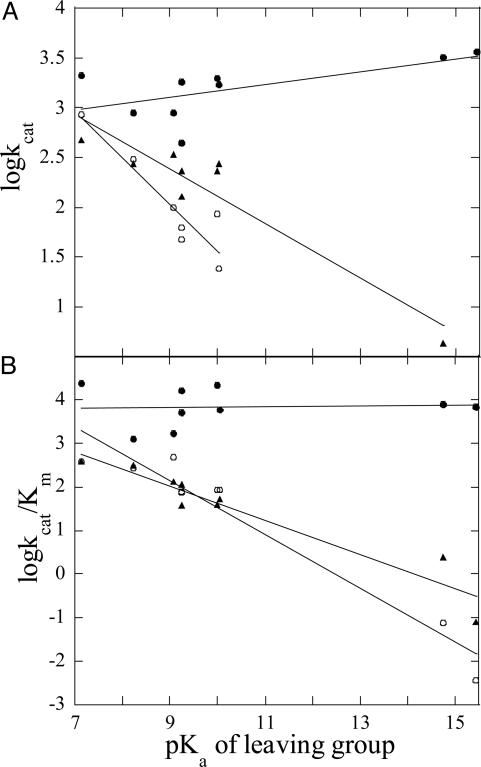
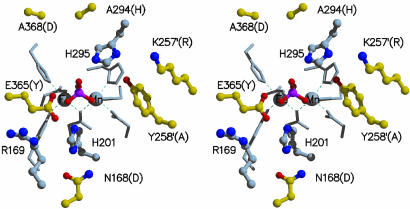
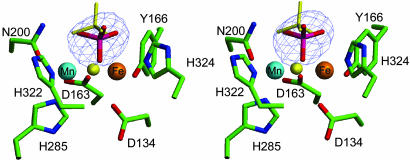
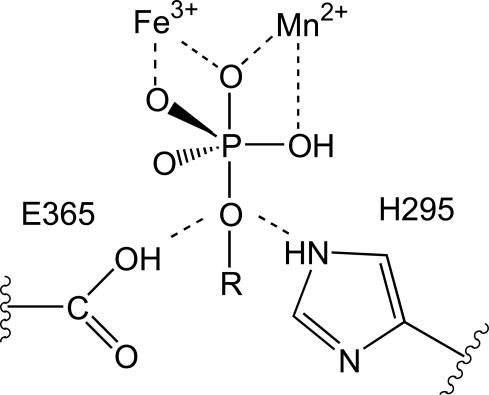
Purple acid phosphatases (PAPs) are a family of binuclear metalloenzymes that catalyze the hydrolysis of phosphoric acid esters and anhydrides. A PAP in sweet potato has a unique, strongly antiferromagnetically coupled Fe(III)-Mn(II) center and is distinguished from other PAPs by its increased catalytic efficiency for a range of activated and unactivated phosphate esters, its strict requirement for Mn(II), and the presence of a mu-oxo bridge at pH 4.90. This enzyme displays maximum catalytic efficiency (k(cat)/K(m)) at pH 4.5, whereas its catalytic rate constant (k(cat)) is maximal at near-neutral pH, and, in contrast to other PAPs, its catalytic parameters are not dependent on the pK(a) of the leaving group. The crystal structure of the phosphate-bound Fe(III)-Mn(II) PAP has been determined to 2.5-A resolution (final R(free) value of 0.256). Structural comparisons of the active site of sweet potato, red kidney bean, and mammalian PAPs show several amino acid substitutions in the sweet potato enzyme that can account for its increased catalytic efficiency. The phosphate molecule binds in an unusual tripodal mode to the two metal ions, with two of the phosphate oxygen atoms binding to Fe(III) and Mn(II), a third oxygen atom bridging the two metal ions, and the fourth oxygen pointing toward the substrate binding pocket. This binding mode is unique among the known structures in this family but is reminiscent of phosphate binding to urease and of sulfate binding to lambda protein phosphatase. The structure and kinetics support the hypothesis that the bridging oxygen atom initiates hydrolysis.
Figures
References
-
- Dismukes, G. C. (1996) Chem. Rev. 96, 2909-2926. - PubMed
-
- Wilcox, D. E. (1996) Chem. Rev. 96, 2435-2458. - PubMed
-
- Schenk, G., Korsinczky, M. L. J., Hume, D. A., Hamilton, S. & de Jersey, J. (2000) Gene 255, 419-424. - PubMed
-
- Ketcham, C. M., Baumach, G. A., Bazer, F. W. & Roberts, R. M. (1985) J. Biol. Chem. 260, 5768-5776. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
