

Nature of the promoter activated by C.PvuII, an unusual regulatory protein conserved among restriction-modification systems
- PMID: 15629920
- PMCID: PMC543531
- DOI: 10.1128/JB.187.2.488-497.2005
Nature of the promoter activated by C.PvuII, an unusual regulatory protein conserved among restriction-modification systems
Abstract
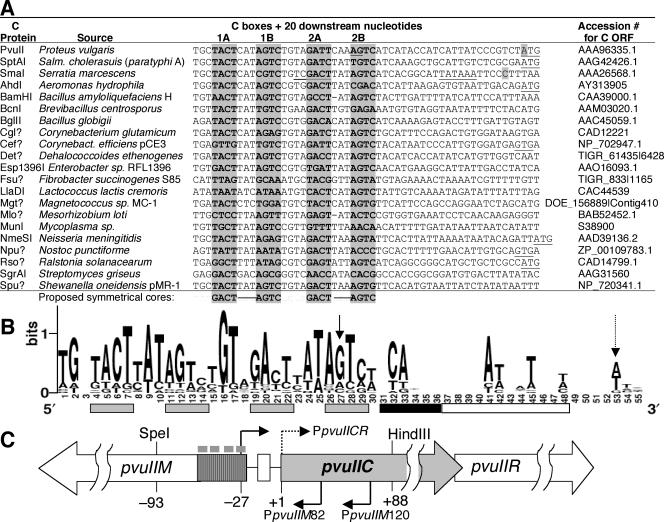
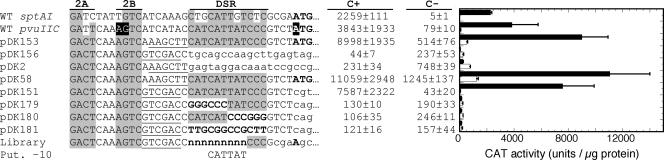
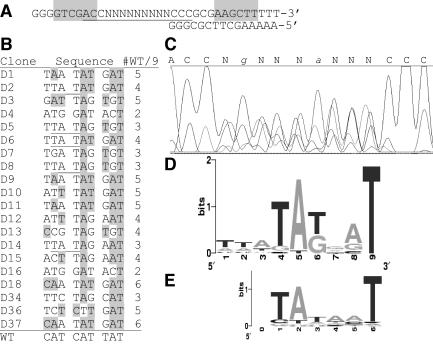
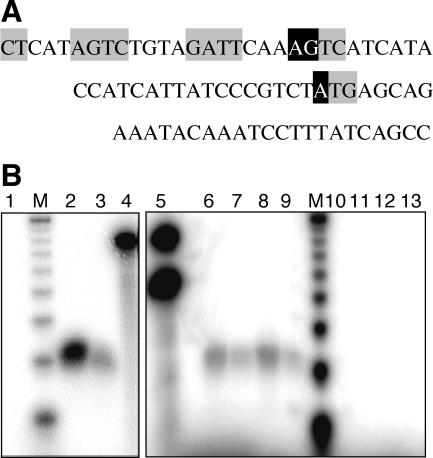
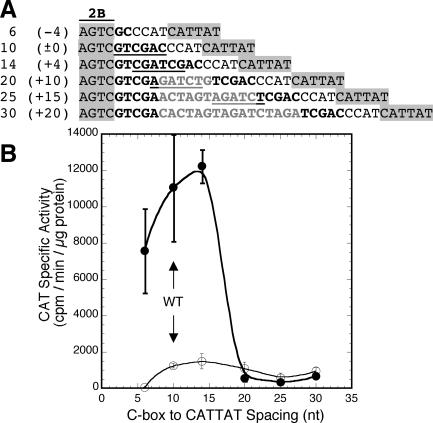
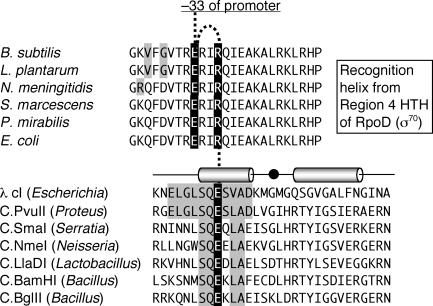
A widely distributed family of small regulators, called C proteins, controls a subset of restriction-modification systems. The C proteins studied to date activate transcription of their own genes and that of downstream endonuclease genes; this arrangement appears to delay endonuclease expression relative to that of the protective methyltransferase when the genes enter a new cell. C proteins bind to conserved sequences called C boxes. In the PvuII system, the C boxes have been reported to extend from -23 to +3 relative to the transcription start for the gene for the C protein, an unexpected starting position relative to a bound activator. This study suggests that transcript initiation within the C boxes represents initial, C-independent transcription of pvuIICR. The major C protein-dependent transcript appears to be a leaderless mRNA starting farther downstream, at the initiation codon for the pvuIIC gene. This conclusion is based on nuclease S1 transcript mapping and the effects of a series of nested deletions in the promoter region. Furthermore, replacing the region upstream of the pvuIIC initiation codon with a library of random oligonucleotides, followed by selection for C-dependent transcription, yielded clones having sequences that resemble -10 promoter hexamers. The -35 hexamer of this promoter would lie within the C boxes. However, the spacing between C boxes/-35 and the apparent -10 hexamer can be varied by +/-4 bp with little effect. This suggests that, like some other activator-dependent promoters, PpvuIICR may not require a -35 hexamer. Features of this transcription activation system suggest explanations for its broad host range.
Figures
Similar articles
-
Translational independence between overlapping genes for a restriction endonuclease and its transcriptional regulator.BMC Mol Biol. 2010 Nov 19;11:87. doi: 10.1186/1471-2199-11-87. BMC Mol Biol. 2010. PMID: 21092102 Free PMC article.
-
Role and mechanism of action of C. PvuII, a regulatory protein conserved among restriction-modification systems.J Bacteriol. 2000 Jan;182(2):477-87. doi: 10.1128/JB.182.2.477-487.2000. J Bacteriol. 2000. PMID: 10629196 Free PMC article.
-
Regulatory circuit based on autogenous activation-repression: roles of C-boxes and spacer sequences in control of the PvuII restriction-modification system.Nucleic Acids Res. 2007;35(20):6935-52. doi: 10.1093/nar/gkm837. Epub 2007 Oct 11. Nucleic Acids Res. 2007. PMID: 17933763 Free PMC article.
-
Regulation of the EcoRI restriction-modification system: Identification of ecoRIM gene promoters and their upstream negative regulators in the ecoRIR gene.Gene. 2007 Oct 1;400(1-2):140-9. doi: 10.1016/j.gene.2007.06.006. Epub 2007 Jun 14. Gene. 2007. PMID: 17618069
-
The TyrR regulon.Mol Microbiol. 2005 Jan;55(1):16-26. doi: 10.1111/j.1365-2958.2004.04385.x. Mol Microbiol. 2005. PMID: 15612913 Review.
Cited by
-
Translational independence between overlapping genes for a restriction endonuclease and its transcriptional regulator.BMC Mol Biol. 2010 Nov 19;11:87. doi: 10.1186/1471-2199-11-87. BMC Mol Biol. 2010. PMID: 21092102 Free PMC article.
-
A novel restriction-modification system is responsible for temperature-dependent phage resistance in Listeria monocytogenes ECII.Appl Environ Microbiol. 2012 Mar;78(6):1995-2004. doi: 10.1128/AEM.07086-11. Epub 2012 Jan 13. Appl Environ Microbiol. 2012. PMID: 22247158 Free PMC article.
-
Natural C-independent expression of restriction endonuclease in a C protein-associated restriction-modification system.Nucleic Acids Res. 2016 Apr 7;44(6):2646-60. doi: 10.1093/nar/gkv1331. Epub 2015 Dec 9. Nucleic Acids Res. 2016. PMID: 26656489 Free PMC article.
-
A bistable hysteretic switch in an activator-repressor regulated restriction-modification system.Nucleic Acids Res. 2013 Jul;41(12):6045-57. doi: 10.1093/nar/gkt324. Epub 2013 Apr 29. Nucleic Acids Res. 2013. PMID: 23630319 Free PMC article.
-
To be or not to be: regulation of restriction-modification systems and other toxin-antitoxin systems.Nucleic Acids Res. 2014 Jan;42(1):70-86. doi: 10.1093/nar/gkt711. Epub 2013 Aug 13. Nucleic Acids Res. 2014. PMID: 23945938 Free PMC article. Review.
References
-
- Anton, B. P., D. F. Heiter, J. S. Benner, E. J. Hess, L. Greenough, L. S. Moran, B. E. Slatko, and J. E. Brooks. 1997. Cloning and characterization of the BglII restriction-modification system reveal a possible evolutionary footprint. Gene 187:19-27. - PubMed
-
- Balbas, P., X. Soberon, E. Merino, M. Zurita, H. Lomeli, F. Valle, N. Flores, and F. Bolivar. 1986. Plasmid vector pBR322 and its special-purpose derivatives—a review. Gene 50:3-40. - PubMed
-
- Bart, A., J. Dankert, and A. van der Ende. 1999. Operator sequences for the regulatory proteins of restriction modification systems. Mol. Microbiol. 31:1277-1278. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
