

Synergy of IL-21 and IL-15 in regulating CD8+ T cell expansion and function
- PMID: 15630141
- PMCID: PMC2212766
- DOI: 10.1084/jem.20041057
Synergy of IL-21 and IL-15 in regulating CD8+ T cell expansion and function
Abstract
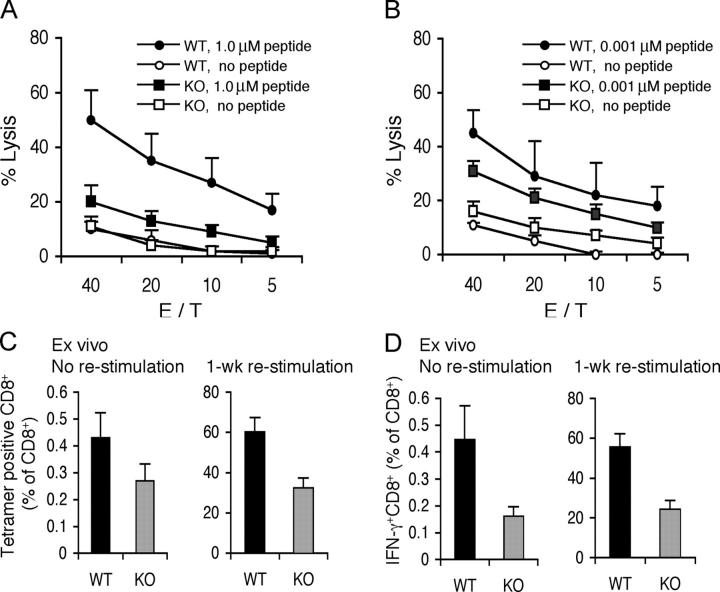
Interleukin (IL)-21 is the most recently recognized of the cytokines that share the common cytokine receptor gamma chain (gamma(c)), which is mutated in humans with X-linked severe combined immunodeficiency. We now report that IL-21 synergistically acts with IL-15 to potently promote the proliferation of both memory (CD44high) and naive (CD44low) phenotype CD8+ T cells and augment interferon-gamma production in vitro. IL-21 also cooperated, albeit more weakly, with IL-7, but not with IL-2. Correspondingly, the expansion and cytotoxicity of CD8+ T cells were impaired in IL-21R-/- mice. Moreover, in vivo administration of IL-21 in combination with IL-15 boosted antigen-specific CD8+ T cell numbers and resulted in a cooperative effect on tumor regression, with apparent cures of large, established B16 melanomas. Thus, our studies reveal that IL-21 potently regulates CD8+ T cell expansion and effector function, primarily in a synergistic context with IL-15.
Figures






References
-
- Noguchi, M., H. Yi, H.M. Rosenblatt, A.H. Filipovich, S. Adelstein, W.S. Modi, O.W. McBride, and W.J. Leonard. 1993. Interleukin-2 receptor gamma chain mutation results in X-linked severe combined immunodeficiency in humans. Cell. 73:147–157. - PubMed
-
- Leonard, W.J. 2001. Cytokines and immunodeficiency diseases. Nat. Rev. Immunol. 1:200–208. - PubMed
-
- Parrish-Novak, J., S.R. Dillon, A. Nelson, A. Hammond, C. Sprecher, J.A. Gross, J. Johnston, K. Madden, W. Xu, J. West, et al. 2000. Interleukin 21 and its receptor are involved in NK cell expansion and regulation of lymphocyte function. Nature. 408:57–63. - PubMed
-
- Kasaian, M.T., M.J. Whitters, L.L. Carter, L.D. Lowe, J.M. Jussif, B. Deng, K.A. Johnson, J.S. Witek, M. Senices, R.F. Konz, et al. 2002. IL-21 limits NK cell responses and promotes antigen-specific T cell activation: a mediator of the transition from innate to adaptive immunity. Immunity. 16:559–569. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
