

Interaction between GATA and the C/EBP family of transcription factors is critical in GATA-mediated suppression of adipocyte differentiation
- PMID: 15632071
- PMCID: PMC543409
- DOI: 10.1128/MCB.25.2.706-715.2005
Interaction between GATA and the C/EBP family of transcription factors is critical in GATA-mediated suppression of adipocyte differentiation
Abstract
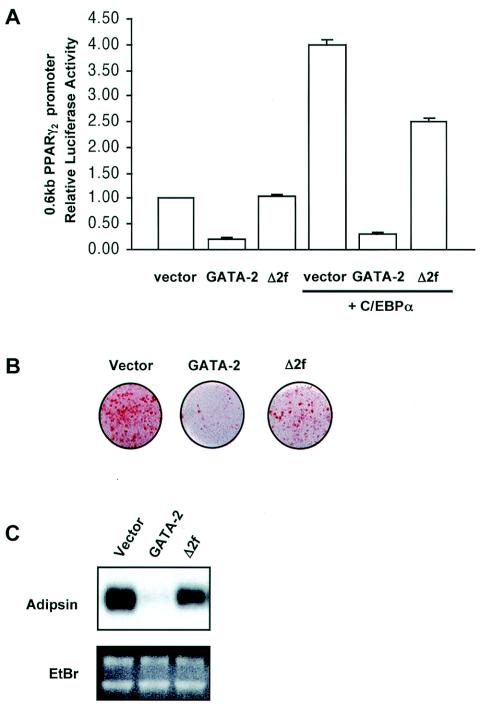
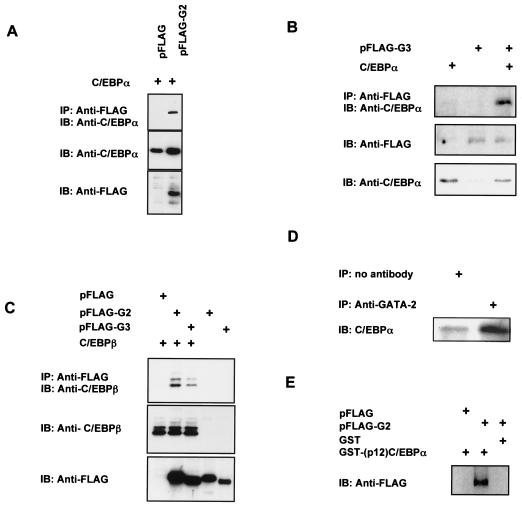
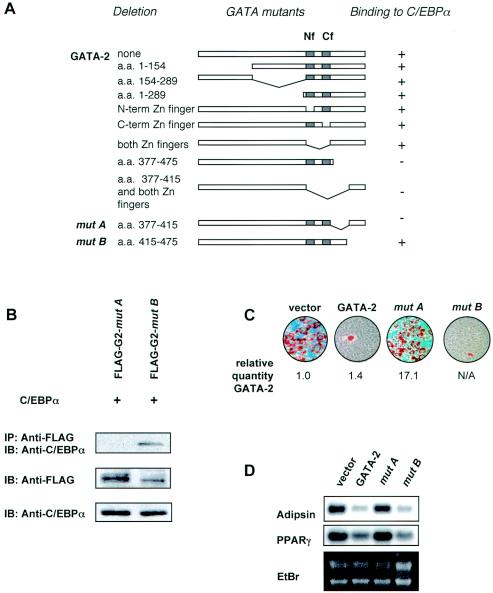
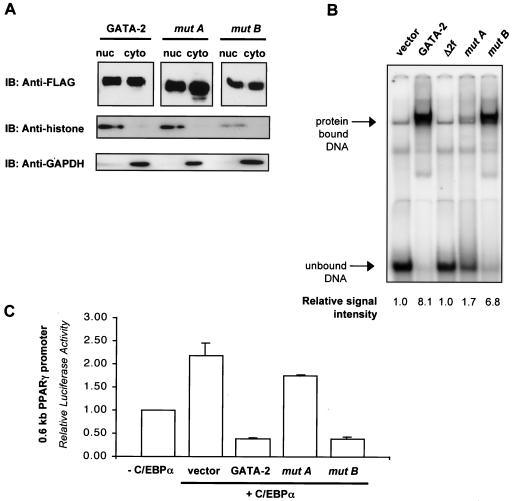
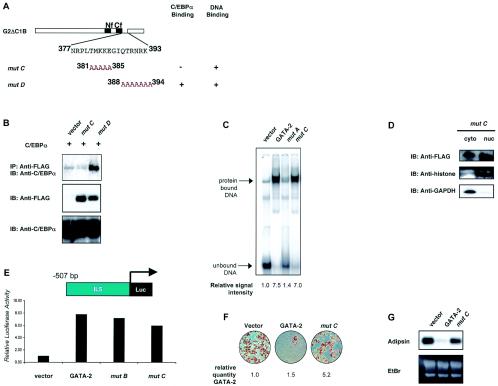
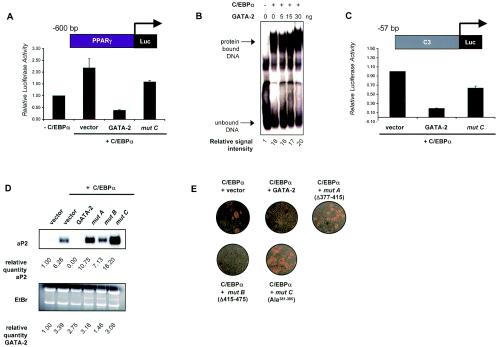
We have previously demonstrated that GATA-2 and GATA-3 are expressed in adipocyte precursors and control the preadipocyte-to-adipocyte transition. Constitutive expression of both GATA-2 and GATA-3 suppressed adipocyte differentiation, partially through direct binding to the peroxisome proliferator-activated receptor gamma (PPARgamma) promoter and suppression of its basal activity. In the present study, we demonstrate that both GATA-2 and GATA-3 form protein complexes with CCAAT/enhancer binding protein alpha (C/EBPalpha) and C/EBPbeta, members of a family of transcription factors that are integral to adipogenesis. We mapped this interaction to the basic leucine zipper domain of C/EBPalpha and a region adjacent to the carboxyl zinc finger of GATA-2. The interaction between GATA and C/EBP factors is critical for the ability of GATA to suppress adipocyte differentiation. Thus, these results show that in addition to its previously recognized function in suppressing PPARgamma transcriptional activity, interaction of GATA factors with C/EBP is necessary for their ability to negatively regulate adipogenesis.
Figures
References
-
- Boyes, J., P. Byfield, Y. Nakatani, and V. Ogryzko. 1998. Regulation of activity of the transcription factor GATA-1 by acetylation. Nature 396:594-598. - PubMed
-
- Cao, Z., R. M. Umek, and S. L. McKnight. 1991. Regulated expression of three C/EBP isoforms during adipose conversion of 3T3-L1 cells. Genes Dev. 5:1538-1552. - PubMed
-
- Chen, P. L., D. J. Riley, Y. Chen, and W. H. Lee. 1996. Retinoblastoma protein positively regulates terminal adipocyte differentiation through direct interaction with C/EBPs. Genes Dev. 10:2794-2804. - PubMed
-
- Crawford, S. E., C. Qi, P. Misra, V. Stellmach, M. S. Rao, J. D. Engel, Y. Zhu, and J. K. Reddy. 2002. Defects of the heart, eye, and megakaryocytes in peroxisome proliferator activator receptor-binding protein (PBP) null embryos implicate GATA family of transcription factors. J. Biol. Chem. 277:3585-3592. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials