

Computational prediction of miRNAs in Arabidopsis thaliana
- PMID: 15632092
- PMCID: PMC540280
- DOI: 10.1101/gr.2908205
Computational prediction of miRNAs in Arabidopsis thaliana
Abstract
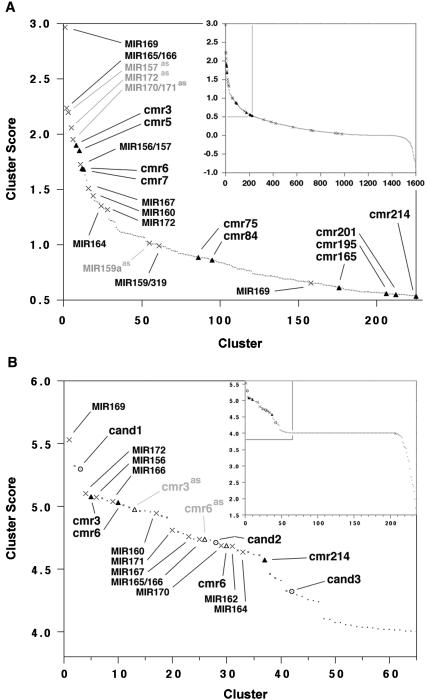
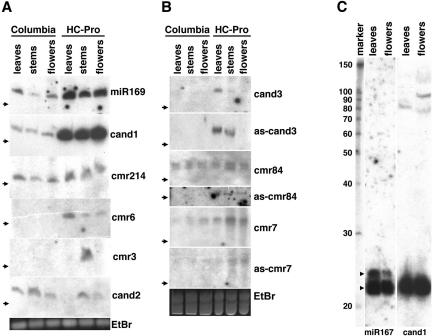
MicroRNAs (miRNAs) are post-transcriptional regulators of gene expression in animals and plants. Comparative genomic computational methods have been developed to predict new miRNAs in worms, flies, and humans. Here, we present a novel single genome approach for the detection of miRNAs in Arabidopsis thaliana. This was initiated by producing a candidate miRNA-target data set using an algorithm called findMiRNA, which predicts potential miRNAs within candidate precursor sequences that have corresponding target sites within transcripts. From this data set, we used a characteristic divergence pattern of miRNA precursor families to select 13 potential new miRNAs for experimental verification, and found that corresponding small RNAs could be detected for at least eight of the candidate miRNAs. Expression of some of these miRNAs appears to be under developmental control. Our results are consistent with the idea that targets of miRNAs encompass a wide range of transcripts, including those for F-box factors, ubiquitin conjugases, Leucine-rich repeat proteins, and metabolic enzymes, and that regulation by miRNAs might be widespread in the genome. The entire set of annotated transcripts in the Arabidopsis genome has been run through find MiRNA to yield a data set that will enable identification of potential miRNAs directed against any target gene.
Figures

References
-
- Bartel, D.P. 2004. MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell 116: 281-297. - PubMed
-
- Bergmann, A. and Lane, M.E. 2003. HIDden targets of microRNAs for growth control. Trends Biochem. Sci. 28: 461-463. - PubMed
-
- Brennecke, J., Hipfner, D.R., Stark, A., Russell, R.B., and Cohen, S.M. 2003. bantam encodes a developmentally regulated microRNA that controls cell proliferation and regulates the proapoptotic gene hid in Drosophila. Cell 113: 25-36. - PubMed
Web site references
-
- http://sundarlab.ucdavis.edu/mirna/; This study.
-
- http://www.arabidopsis.org; The Arabidopsis Information resource (TAIR).
-
- http://www.sanger.ac.uk/Software/Rfam/mirna/; The miRNA Registry.
-
- http://www.gramene.org/; Gramene: A comparative mapping resource for grains.
-
- http://gac.bcc.orst.edu/smallRNA/; Arabidopsis thaliana Small RNA project.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases