

Identification of C7orf11 (TTDN1) gene mutations and genetic heterogeneity in nonphotosensitive trichothiodystrophy
- PMID: 15645389
- PMCID: PMC1196401
- DOI: 10.1086/428141
Identification of C7orf11 (TTDN1) gene mutations and genetic heterogeneity in nonphotosensitive trichothiodystrophy
Abstract
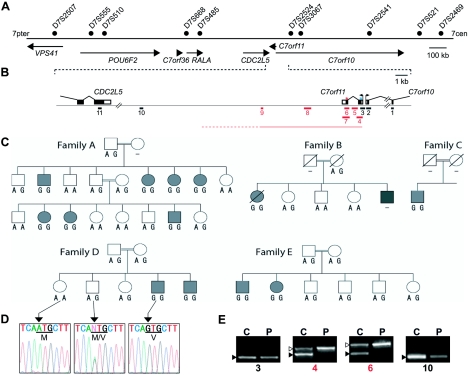
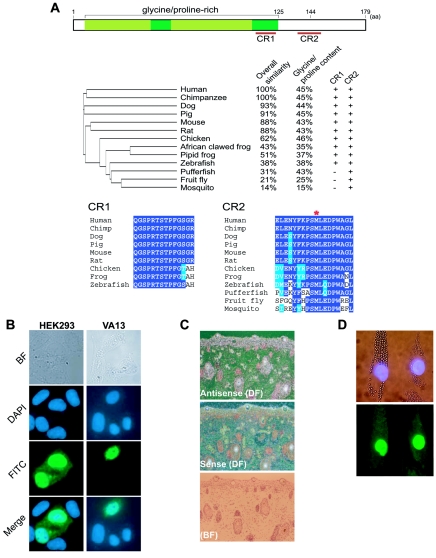
We have identified C7orf11, which localizes to the nucleus and is expressed in fetal hair follicles, as the first disease gene for nonphotosensitive trichothiodystrophy (TTD). C7orf11 maps to chromosome 7p14, and the disease locus has been designated "TTDN1" (TTD nonphotosensitive 1). Mutations were found in patients with Amish brittle-hair syndrome and in other nonphotosensititive TTD cases with mental retardation and decreased fertility but not in patients with Sabinas syndrome or Pollitt syndrome. Therefore, genetic heterogeneity in nonphotosensitive TTD is a feature similar to that observed in photosensitive TTD, which is caused by mutations in transcription factor II H (TFIIH) subunit genes. Comparative immunofluorescence analysis, however, suggests that C7orf11 does not influence TFIIH directly. Given the absence of cutaneous photosensitivity in the patients with C7orf11 mutations, together with the protein's nuclear localization, C7orf11 may be involved in transcription but not DNA repair.
Figures
References
Electronic-Database Information
-
- The Chromosome 7 Annotation Project, http://www.chr7.org/ (for seven candidate genes on 7p14)
-
- Ensembl, http://www.ensembl.org/ (for proteins fromchimpanzee [accession number ENSPTRP00000032652],rat [accession number ENSRNOP00000018746], and chicken [accession number ENSGALP00000020100])
-
- GenBank, http://www.ncbi.nlm.nih.gov/Genbank/ (forhuman C7orf11 mRNA [accession number NM_138701] and cDNA sequences from zebrafish [accession number BC062385] and pig [accession number BP456435])
-
- NCBI, http://www.ncbi.nlm.nih.gov/ (for human C7orf11 [accession number NP_619646] and orthologues from mouse [accession number BAB27916], frog [accession number NP_989025], pufferfish [accession number CAF91712], fruitfly [accession number NP_648690], and mosquito [accession number XP_318005])
-
- Online Mendelian Inheritance in Man (OMIM), http://www.ncbi.nlm.nih.gov/Omim/ (for Amish brittle-hair syndrome, Sabinas syndrome, and Pollitt syndrome)
References
-
- Alfandari S, Delaporte E, van Neste D, Lucidarme-Delespierre E, Piette F, Bergoend H (1993) A new case of isolated trichothiodystrophy. Dermatology 186:197–200 - PubMed
-
- Botta E, Nardo T, Broughton BC, Marinoni S, Lehmann AR, Stefanini M (1998) Analysis of mutations in the XPD gene in Italian patients with trichothiodystrophy: site of mutation correlates with repair deficiency, but gene dosage appears to determine clinical severity. Am J Hum Genet 63:1036–1048 - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
