

Organelle-specific control of intracellular transport: distinctly targeted isoforms of the regulator Klar
- PMID: 15647372
- PMCID: PMC551502
- DOI: 10.1091/mbc.e04-10-0920
Organelle-specific control of intracellular transport: distinctly targeted isoforms of the regulator Klar
Abstract
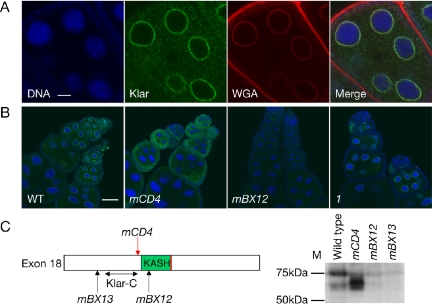
Microtubule-based transport in cells is powered by a small set of distinct motors, yet timing and destination of transport can be controlled in a cargo-specific manner. The mechanistic basis for this specificity is not understood. To address this question, we analyzed the Drosophila Klarsicht (Klar) protein that regulates distinct microtubule-based transport processes. We find that localization of Klar to its cargoes is crucial for Klar function. Using mutations, we identify functionally important regions of Klar that confer distinct cargo specificity. In ovaries, Klar is present on the nuclear envelope, a localization that requires the C-terminal KASH domain. In early embryos, Klar is attached to lipid droplets, a localization mediated by a novel C-terminal domain encoded by an alternatively spliced exon. In cultured cells, these two domains are sufficient for targeting to the correct intracellular location. Our analysis disentangles Klar's modular organization: we propose that a core region integral to motor regulation is attached to variable domains so that the cell can target regulators with overlapping, yet distinct functions to specific cargoes. Such isoform variation may be a general strategy for adapting a common regulatory mechanism to specifically control motion and positioning of multiple organelles.
Figures






Similar articles
-
Drosophila KASH-domain protein Klarsicht regulates microtubule stability and integrin receptor localization during collective cell migration.Dev Biol. 2015 Nov 1;407(1):103-14. doi: 10.1016/j.ydbio.2015.08.003. Epub 2015 Aug 3. Dev Biol. 2015. PMID: 26247519 Free PMC article.
-
Targeting the motor regulator Klar to lipid droplets.BMC Cell Biol. 2011 Feb 24;12:9. doi: 10.1186/1471-2121-12-9. BMC Cell Biol. 2011. PMID: 21349165 Free PMC article.
-
Drosophila klarsicht has distinct subcellular localization domains for nuclear envelope and microtubule localization in the eye.Genetics. 2004 Nov;168(3):1385-93. doi: 10.1534/genetics.104.028662. Genetics. 2004. PMID: 15579692 Free PMC article.
-
Bidirectional transport along microtubules.Curr Biol. 2004 Jul 13;14(13):R525-37. doi: 10.1016/j.cub.2004.06.045. Curr Biol. 2004. PMID: 15242636 Review.
-
Vesicle transport: klarsicht clears up the matter.Curr Biol. 1998 Jul 16;8(15):R542-4. doi: 10.1016/s0960-9822(07)00343-0. Curr Biol. 1998. PMID: 9705929 Review.
Cited by
-
UNC-83 IS a KASH protein required for nuclear migration and is recruited to the outer nuclear membrane by a physical interaction with the SUN protein UNC-84.Mol Biol Cell. 2006 Apr;17(4):1790-801. doi: 10.1091/mbc.e05-09-0894. Epub 2006 Feb 15. Mol Biol Cell. 2006. PMID: 16481402 Free PMC article.
-
PAT proteins, an ancient family of lipid droplet proteins that regulate cellular lipid stores.Biochim Biophys Acta. 2009 Jun;1791(6):419-40. doi: 10.1016/j.bbalip.2009.04.002. Epub 2009 Apr 16. Biochim Biophys Acta. 2009. PMID: 19375517 Free PMC article. Review.
-
Drosophila KASH-domain protein Klarsicht regulates microtubule stability and integrin receptor localization during collective cell migration.Dev Biol. 2015 Nov 1;407(1):103-14. doi: 10.1016/j.ydbio.2015.08.003. Epub 2015 Aug 3. Dev Biol. 2015. PMID: 26247519 Free PMC article.
-
In-vivo centrifugation of Drosophila embryos.J Vis Exp. 2010 Jun 23;(40):2005. doi: 10.3791/2005. J Vis Exp. 2010. PMID: 20613707 Free PMC article.
-
Lipid droplet motility and organelle contacts.Contact (Thousand Oaks). 2019 Jan-Dec;2:10.1177/2515256419895688. doi: 10.1177/2515256419895688. Epub 2019 Dec 16. Contact (Thousand Oaks). 2019. PMID: 31909374 Free PMC article.
References
-
- Almenar-Queralt, A., and Goldstein, L. S. (2001). Linkers, packages and pathways: new concepts in axonal transport. Curr. Opin. Neurobiol. 11, 550-557. - PubMed
-
- Apel, E. D., Lewis, R. M., Grady, R. M., and Sanes, J. R. (2000). Syne-1, a dystrophin- and Klarsicht-related protein associated with synaptic nuclei at the neuromuscular junction. J. Biol. Chem. 275, 31986-31995. - PubMed
-
- Brown, D. (2001). Lipid droplets: proteins floating on a pool of fat. Curr. Biol. 11, R446-R449. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
