

Review
doi: 10.1016/j.pbiomolbio.2004.07.006.
Epub 2004 Oct 7.
Membrane protein crystallization in amphiphile phases: practical and theoretical considerations
Affiliations
- PMID: 15652249
- PMCID: PMC2748814
- DOI: 10.1016/j.pbiomolbio.2004.07.006
Item in Clipboard
Review
Membrane protein crystallization in amphiphile phases: practical and theoretical considerations
Prog Biophys Mol Biol.
2005 Jul.
Abstract
Integral membrane proteins are amphiphilic molecules. In order to enable chromatographic purification and crystallization, a complementary amphiphilic microenvironment must be created and maintained. Various types of amphiphilic phases have been employed in crystallizations and intricate amphiphilic microenvironmental structures have resulted from these and are found inside membrane protein crystals. In this review the process of crystallization is put into the context of amphiphile phase transitions. Finally, practical factors are considered and a pragmatic way is suggested to pursue membrane protein crystallization trials.
Figures
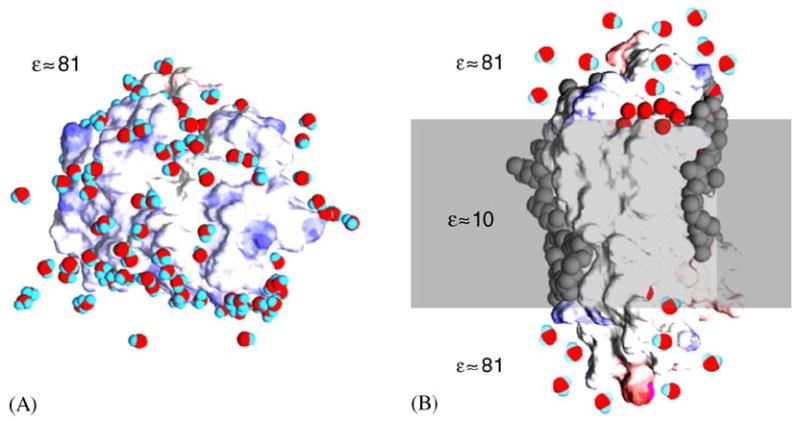
Schematic depiction of a soluble protein and a transmembrane protein in their native environment, an aqueous solution and a membrane bilayer, respectively. Both models are based on high-resolution X-ray crystallographic experimental structures showing protein, water and lipids. Water molecules are red and blue, the surface of the protein is blue where there is negative charge and red where there is positive charge, hydrophobic core and lipids are colored gray. (A) Soluble protein particles are dissolved in a homogenous medium with a dielectric constant ε close to that of distilled water. On their surface they interact with water molecules and with ions. (B) Integral protein particles are dissolved a low dielectric amphipilic medium, the lipid bilayer membrane, contacting the hydrophobic core, while their hydrophilic caps are exposed to an aqueous medium similar to that shown in A for soluble proteins.
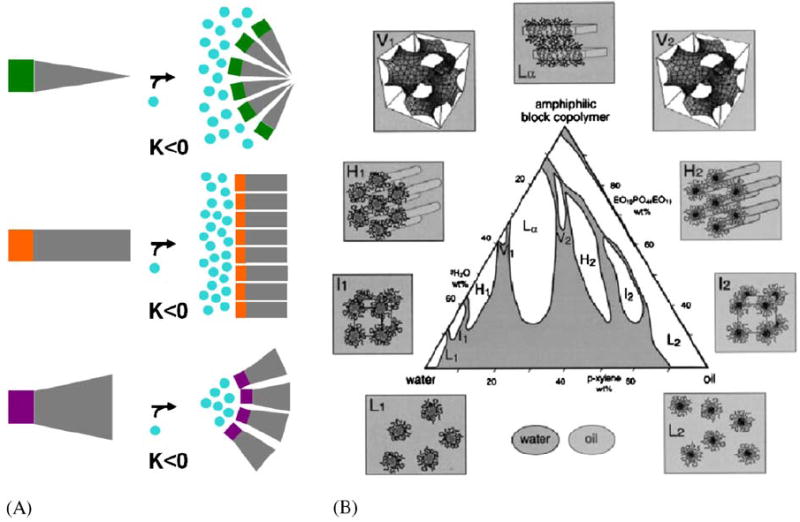
Introduction to small molecule amphiphile classes and an example for amphipile polymorphism. (A) According to the ‘intrinsic curvature hypothesis’ (Gruner, 1985) amphiphiles form supramolecular structures in water due to the hydrophobic effect (Tanford, 1980). In this simplistic view the shape, more specifically the ratio of cross section of hydrophilic headgroup and hydrophobic tail, determines the type of structure they self-assemble into. I.e. wedge-shaped amphiphiles assemble into structures with positive Gaussian curvature K such as micelles, cylinder-shaped amphiphiles assemble into structures with zero Gaussian curvature K such as planar lipid bilayers and, cone-shaped amphiphiles assemble into structures with negative Gaussian curvature K such as bicontinuous cubic phases. (B) Amphiphiles may adopt a multitude of such supramolecular assemblies, macroscopic phases when mixed with water. For a particular three component system, oil, water, amphiphilic block copolymer, Alexandris et al. (1998) describe a record nine different isothermal phases including four cubic, two hexagonal and one lamellar lyotropic liquid crystalline phase and two micellar solutions.
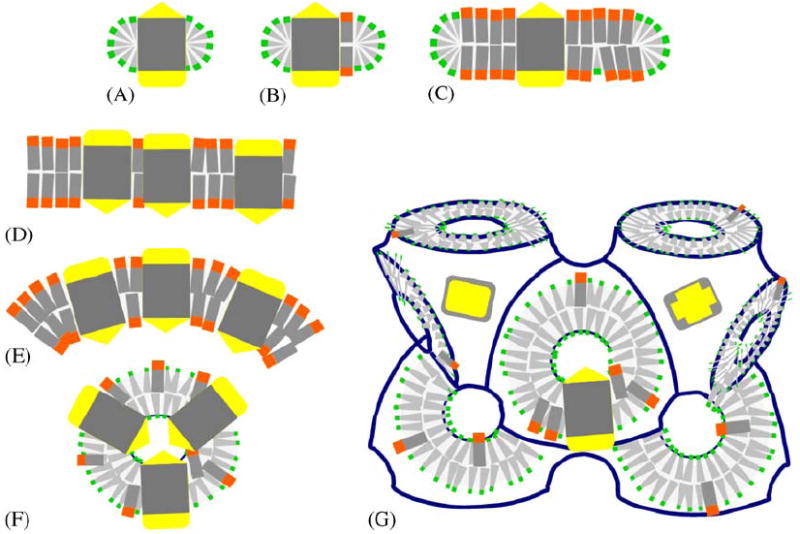
Schematic depiction of different amphiphilic micro-environments that membrane proteins have been incorporated into for crystallization purposes. Membrane proteins are shown with two hydrophilic caps (yellow) and a hydrophobic core (gray), and lipids (orange head) and detergents (green head) provide discrete (A, B, C) or continous (D, E, F, G) amphiphilic assemblies. (A) Protein detergent micelle; (B) Protein detergent complex (PDC); (C) Bicelle protein complex; (D) Planar membrane bilayer with embedded protein; (E) Curved proteoliposome; (F) High-curvature regular spherical shell assembly; (G) Membrane proteins reconstituted in bicontinuous curved membranes of a lipidic cubic phase of the diamond type. In all of these structures the amphiphile molecules are highly mobile (Zulauf, 1991) and thus constitute a ‘fluid’ hydrophobic host medium in which the integral membrane protein is embedded.
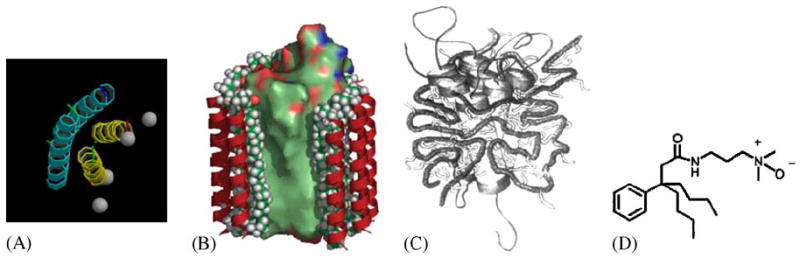
Cartoons of recently developed new classes of amphiphile molecules. (A) Peptitergents are designed peptides that form amphiphatic α-helices. Their hydrophobic faces were made to interact with the hydrophobic face of integral membrane proteins (Schafmeister et al., 1993). (B) Lipopeptides consist of a peptide scaffold supporting two alkyl chains each anchored at the end of an α-helix. They were designed to provide a rigid outer hydrophilic shell surrounding an inner ‘soft’ lipidic core (McGregor, 2003). (C) Amphiphols are amphiphilic polymers with a hydrophilic backbone and hydrophobic grafted chains. They exhibit favorable phase transitions that are expected to be useful for membrane protein crystallization experiments (Tribet et al., 1996). (D) The design of tripod amphiphiles such as #8 in Yu et al. (2000) is based on the rationale that detergents with diminished flexibility should be more prone to form ordered crystal lattices. The core is a terasubstituted carbon atom that limits the flexibility of three attached hydrophobic and a hydrophilic substitutent.
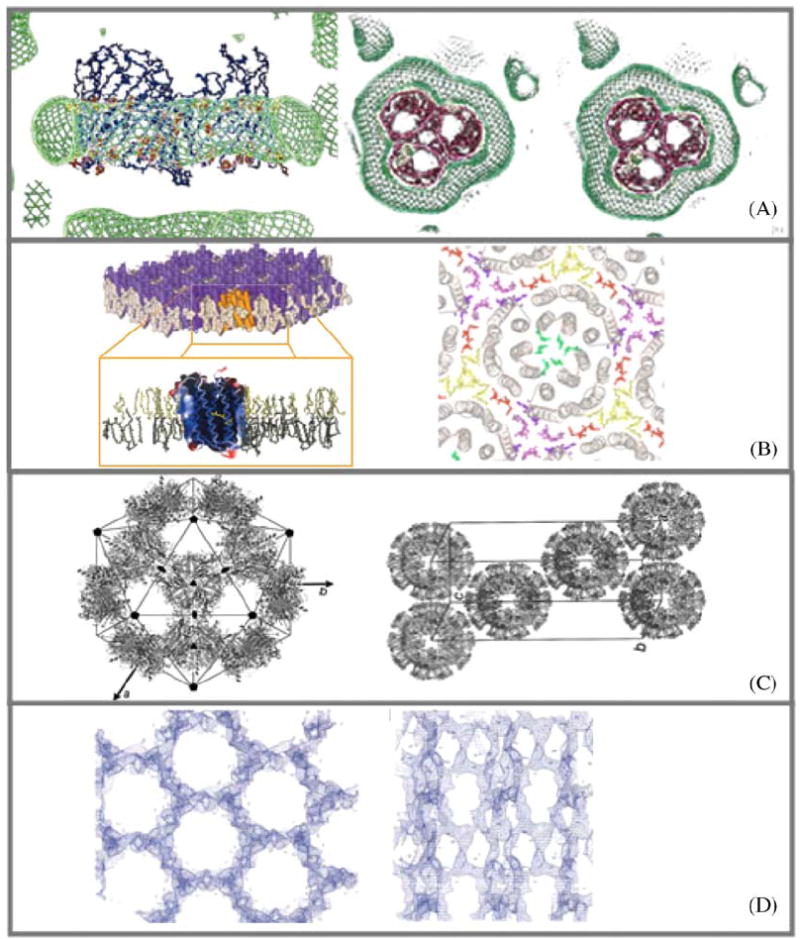
Samples of experimentally determined and deduced amphiphile arrangements in crystals of membrane proteins. (A) ‘Type II’ discontinuous arrangement of micellar Octyl-POE around the hydrophobic perimeter of the detergent C8E4 and OmpF porin (E. coli) R3 crystal (Pebay-Peyroula et al., 1995). Protein detergent micelle with a belt of detergent as shown by neutron density (green). (B) ‘Type I’ continuous arrangement of stacked layers consisting of extended two-dimensional sheets of purple membrane (Halobacterium Salinarum). Bacteriorhodopsin trimers are separated in plane by a belt of native lipids (Belrhali et al., 1999). (C) Discontinuous icosahedral packing of spherical LHC-II proteoliposomes (Liu et al., 2004). (D) Continuous network of β-Octylglucoside extending throughout the entire P3121 crystal of phospholipase A OmplA (E. coli) (Snijder et al., 2003).
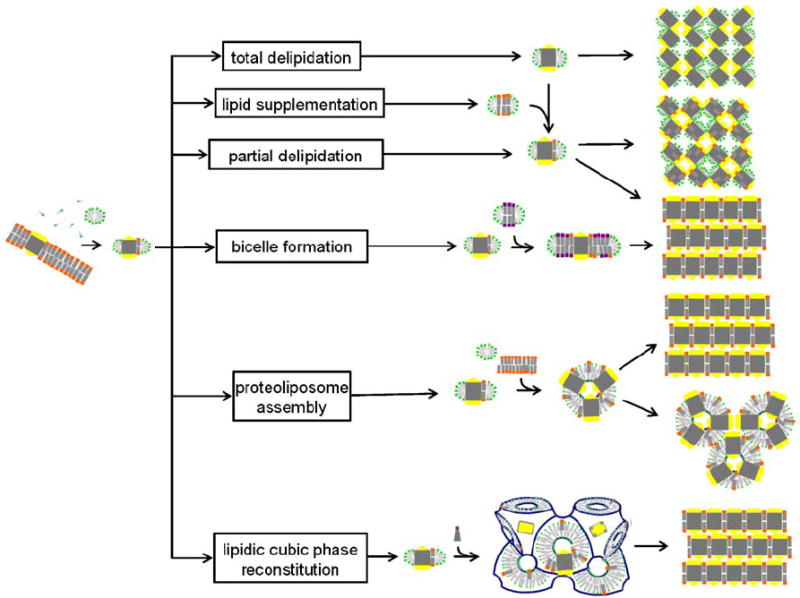
Schematic representation of membrane protein crystallization concepts, routes and examples for the resulting packing arrangements. The detergent-solubilized membrane protein may be delipidated (Pautsch and Schulz, 1998) or may carry affine lipids throughout the purification process. Alternatively, membrane-forming lipids may be added to form a PDC, a protein/detergent/lipid complex (Zhang et al., 2003) or bicelles (Faham and Bowie, 2002). Structured amphiphilic phases can form when lipids with zero or negative spontaneous curvature are added to detergent solubilized membrane protein preparations. Proteoliposomes may fuse and form layered stacks or they may directly assemble into a regular array. Bicontinuous lipidic cubic phases can provide a matrix for embedding membrane proteins and allow these to nucleate and form layered crystals (Nollert et al., 2001).
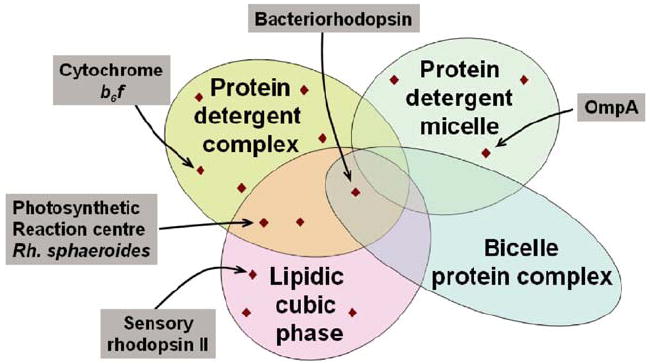
Venn diagram schematizing crystallization method space. Depicted are four different amphiphilic micro-environments that support membrane protein crystallization. For illustration purposes representative examples are pointed out.
Similar articles
-
Widening the protein crystallization bottleneck.Nat Methods. 2006 Dec;3(12):961. doi: 10.1038/nmeth1206-961. Nat Methods. 2006. PMID: 17190005
-
Protein aggregation in silico.Trends Biotechnol. 2007 Jun;25(6):254-61. doi: 10.1016/j.tibtech.2007.03.011. Epub 2007 Apr 12. Trends Biotechnol. 2007. PMID: 17433843 Free PMC article. Review.
-
A model for enhanced nucleation of protein crystals on a fractal porous substrate.Biophys J. 2006 Nov 15;91(10):3857-63. doi: 10.1529/biophysj.106.082545. Epub 2006 Aug 18. Biophys J. 2006. PMID: 16920829 Free PMC article.
-
Pattern formation and fluctuation-induced transitions in protein crystallization.J Chem Phys. 2004 Apr 22;120(16):7708-19. doi: 10.1063/1.1687339. J Chem Phys. 2004. PMID: 15267682
-
Protein crystallization: from purified protein to diffraction-quality crystal.Nat Methods. 2008 Feb;5(2):147-53. doi: 10.1038/nmeth.f.203. Nat Methods. 2008. PMID: 18235435 Review.
Cited by
-
Frequencies of hydrophobic and hydrophilic runs and alternations in proteins of known structure.Protein Sci. 2006 Jan;15(1):102-12. doi: 10.1110/ps.051741806. Protein Sci. 2006. PMID: 16373477 Free PMC article.
-
Orientation and motion of tryptophan interfacial anchors in membrane-spanning peptides.Biochemistry. 2007 Jun 26;46(25):7514-24. doi: 10.1021/bi700082v. Epub 2007 May 27. Biochemistry. 2007. PMID: 17530863 Free PMC article.
-
The Fluidity of Phosphocholine and Maltoside Micelles and the Effect of CHAPS.Biophys J. 2019 May 7;116(9):1682-1691. doi: 10.1016/j.bpj.2019.03.019. Epub 2019 Mar 30. Biophys J. 2019. PMID: 31023535 Free PMC article.
-
Crystallographic characterization of N-oxide tripod amphiphiles.J Am Chem Soc. 2010 Feb 17;132(6):1953-9. doi: 10.1021/ja9085148. J Am Chem Soc. 2010. PMID: 20095541 Free PMC article.
-
Tripod Amphiphiles for Membrane Protein Manipulation.Mol Biosyst. 2010;6:89-94. doi: 10.1039/b915162c. Mol Biosyst. 2010. PMID: 23814603 Free PMC article.
References
-
- Alexandridis P, Olsson U, Lindman B. A record nine different phases (four cubic, two hexagonal, and one lamellar lyotropic liquid crystalline and two micellar solutions) in a ternary isothermal system of an amphiphilic block copolymer and selective solvents (water and oil) Langmuir. 1998;14:2627–2638.
-
- Belrhali H, Nollert P, Royant A, Menzel C, Rosenbusch J, Landau EM, Pebay-Peyroula E. Protein, lipid and water organization in bacteriorhodopsin crystals: a molecular view of the purple membrane at 1.9 angstrom resolution. Structure. 1999;7:909–917. - PubMed
-
- Buchanan SK. β-Barrel proteins in bacterial outer membranes: structure, function and refolding. Curr Opin Struct Biol. 1999;9:455–461. - PubMed
-
- Caffrey M. Membrane protein crystallization. J Struct Biol. 2003;142:108–132. - PubMed
-
- Cartailler J-P, Luecke H. X-ray crystallographic analysis of lipid-protein interactions in the bacteriorhodopsin purple membrane. Ann Rev Biophys Biomol Struct. 2003;32:285–310. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
