

Genetic control of temperature preference in the nematode Caenorhabditis elegans
- PMID: 15654086
- PMCID: PMC1449549
- DOI: 10.1534/genetics.104.036111
Genetic control of temperature preference in the nematode Caenorhabditis elegans
Abstract
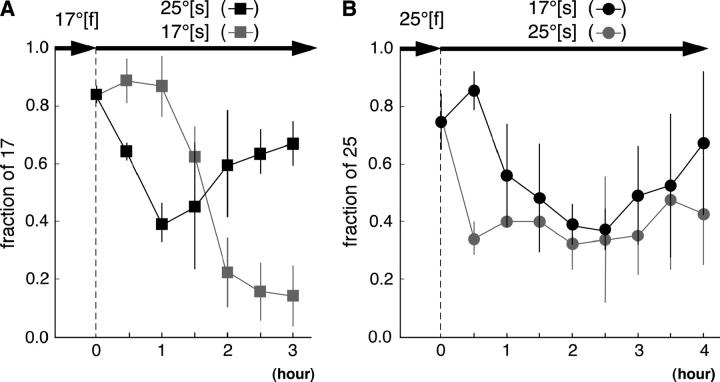
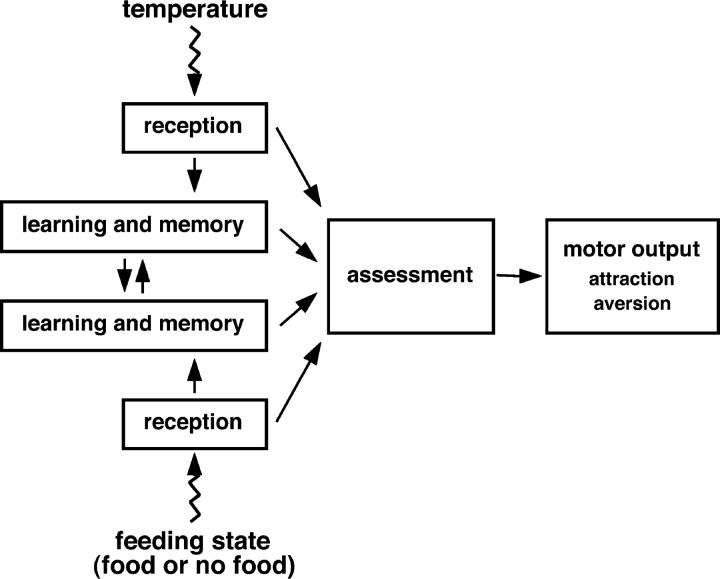
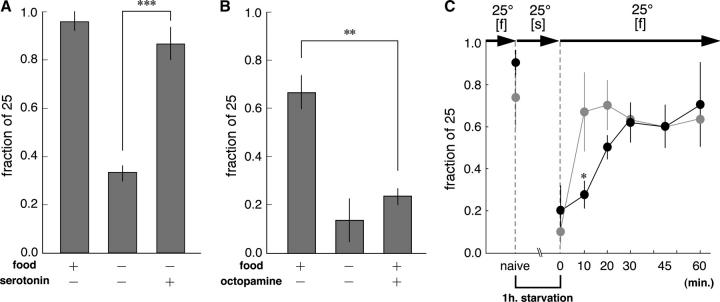
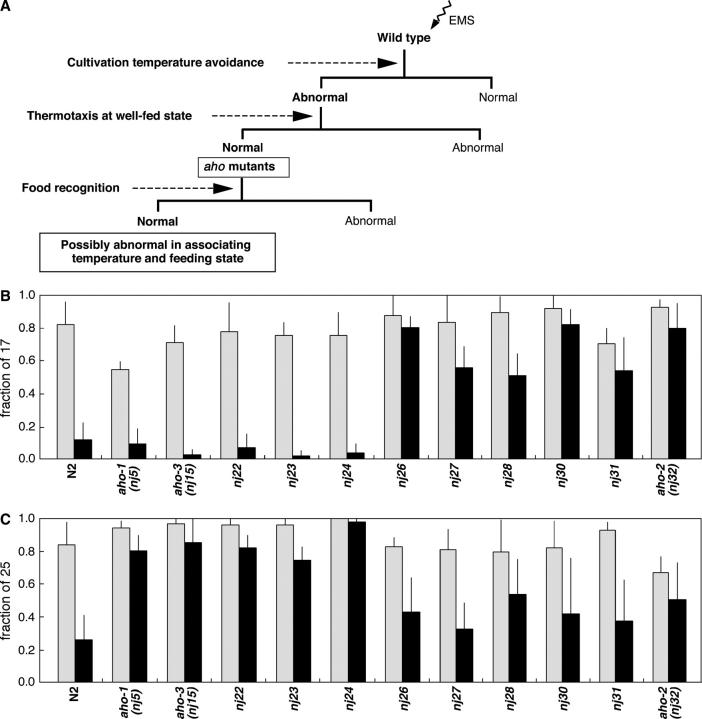
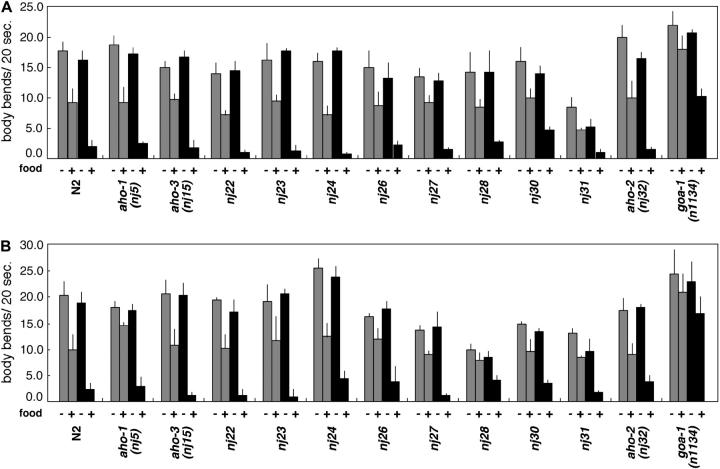
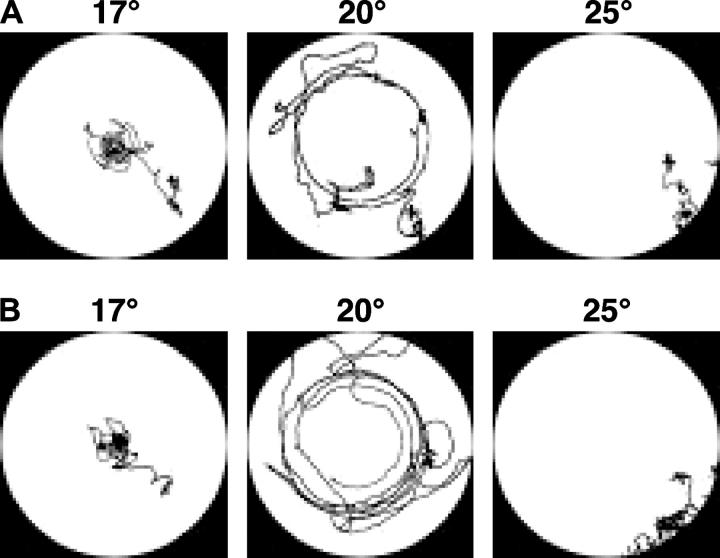
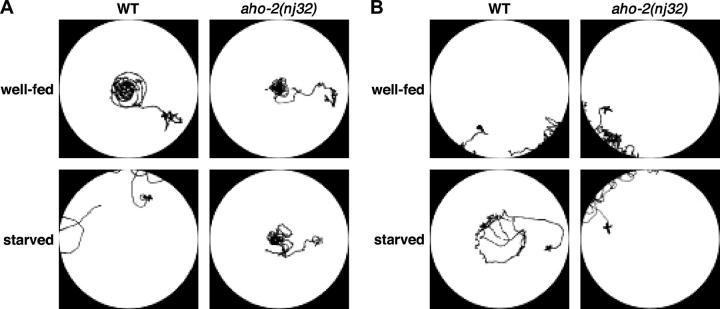
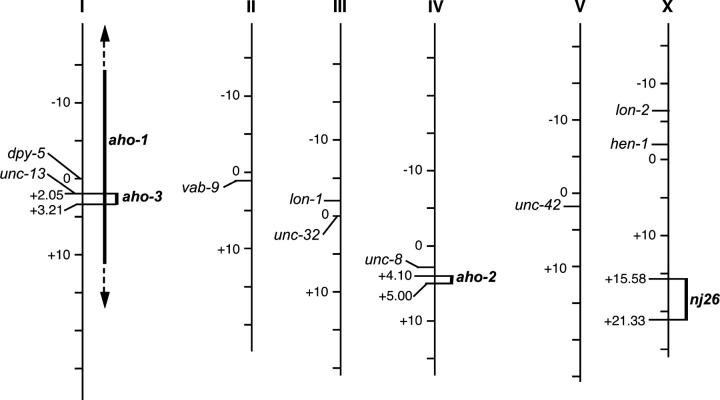
Animals modify behavioral outputs in response to environmental changes. C. elegans exhibits thermotaxis, where well-fed animals show attraction to their cultivation temperature on a thermal gradient without food. We show here that feeding-state-dependent modulation of thermotaxis is a powerful behavioral paradigm for elucidating the mechanism underlying neural plasticity, learning, and memory in higher animals. Starved experience alone could induce aversive response to cultivation temperature. Changing both cultivation temperature and feeding state simultaneously evoked transient attraction to or aversion to the previous cultivation temperature: recultivation of starved animals with food immediately induced attraction to the temperature associated with starvation, although the animals eventually exhibited thermotaxis to the new temperature associated with food. These results suggest that the change in feeding state quickly stimulates the switch between attraction and aversion for the temperature in memory and that the acquisition of new temperature memory establishes more slowly. We isolated aho (abnormal hunger orientation) mutants that are defective in starvation-induced cultivation-temperature avoidance. Some aho mutants responded normally to changes in feeding state with respect to locomotory activity, implying that the primary thermosensation followed by temperature memory formation remains normal and the modulatory aspect of thermotaxis is specifically impaired in these mutants.
Figures
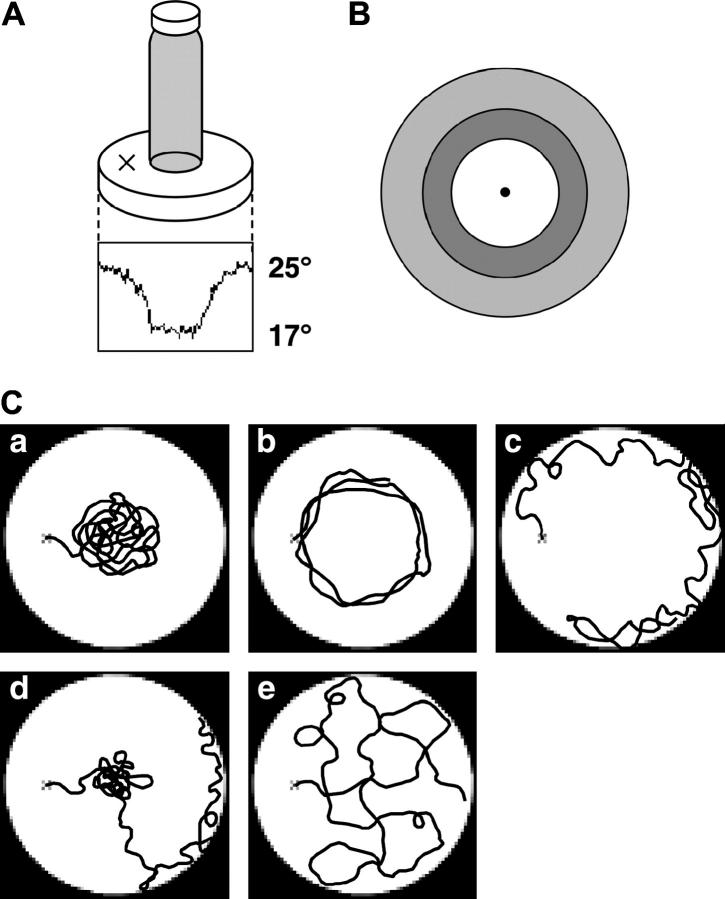
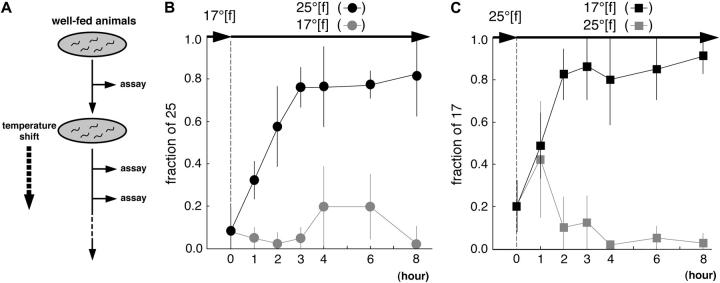
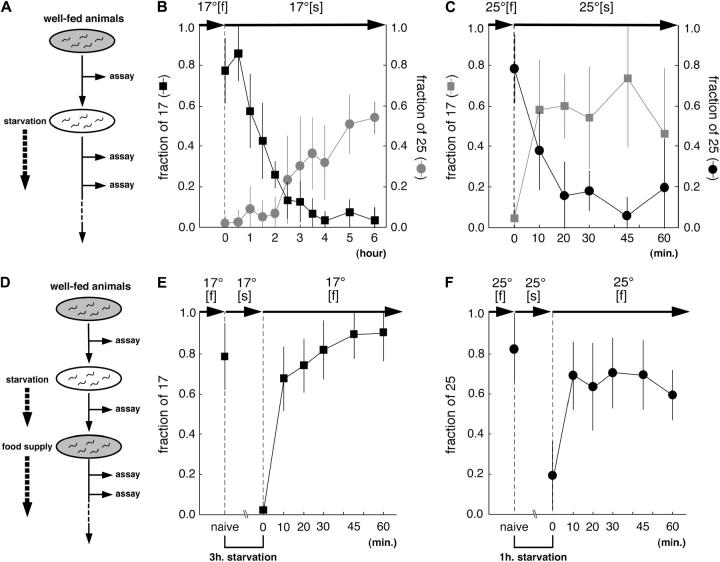
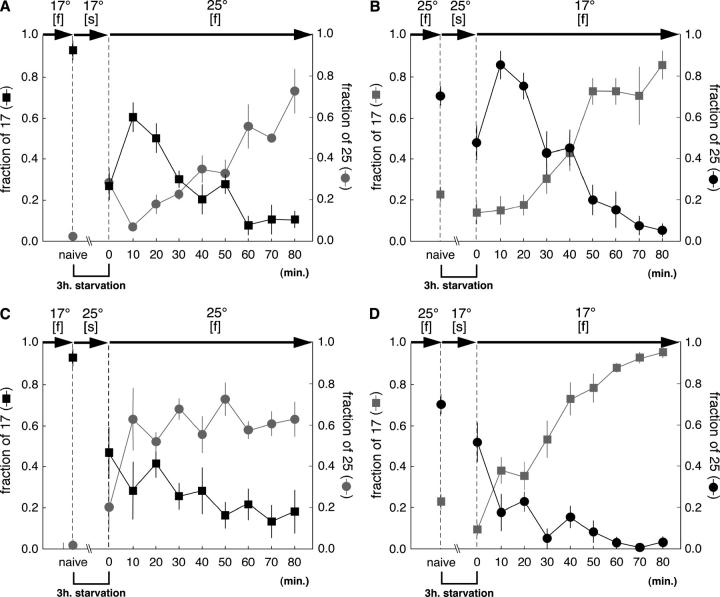
References
-
- Avery, L., and H. R. Horvitz, 1990. Effects of starvation and neuroactive drugs on feeding in Caenorhabditis elegans. J. Exp. Zool. 253: 263–270. - PubMed
-
- Bargmann, C. I., 1998. Neurobiology of the Caenorhabditis elegans genome. Science 282: 2028–2033. - PubMed
-
- Colbert, H. A., and C. I. Bargmann, 1995. Odorant-specific adaptation pathways generate olfactory plasticity in C. elegans. Neuron 14: 803–812. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
