

Ca(2+)- and volume-sensitive chloride currents are differentially regulated by agonists and store-operated Ca2+ entry
- PMID: 15657298
- PMCID: PMC2217498
- DOI: 10.1085/jgp.200409161
Ca(2+)- and volume-sensitive chloride currents are differentially regulated by agonists and store-operated Ca2+ entry
Abstract
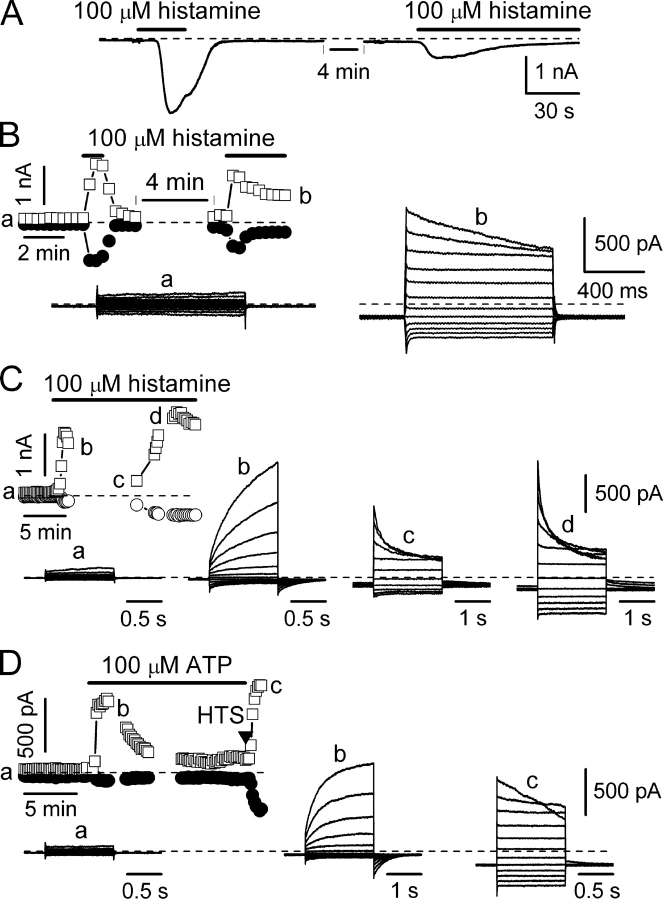
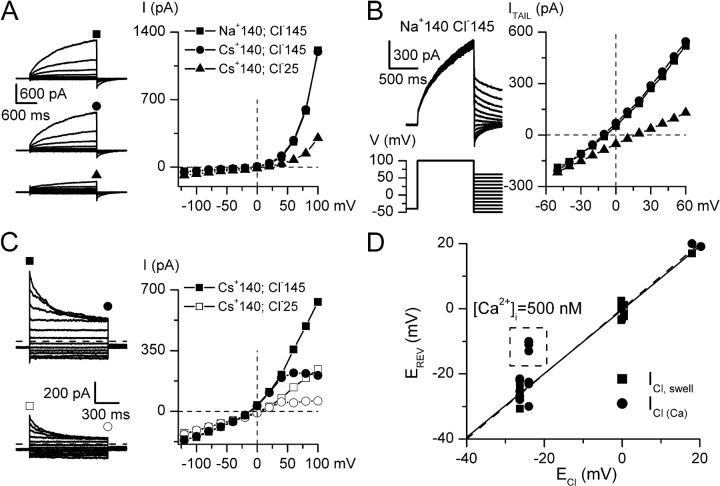
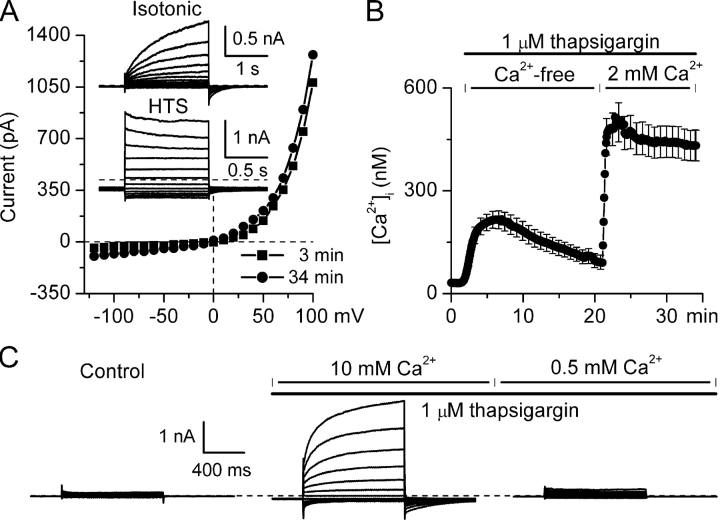
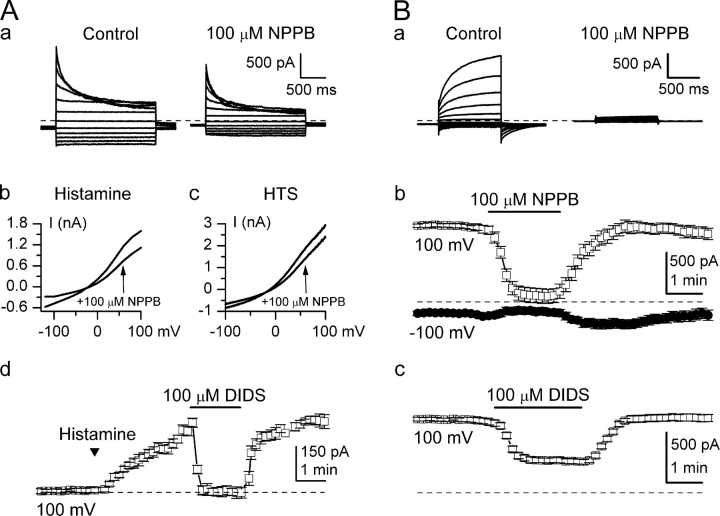
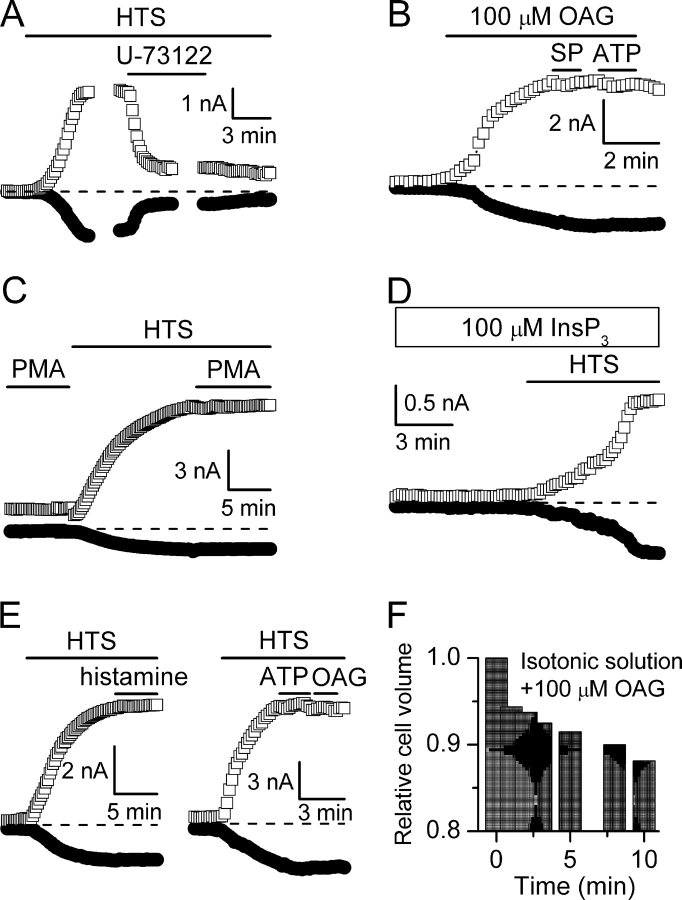
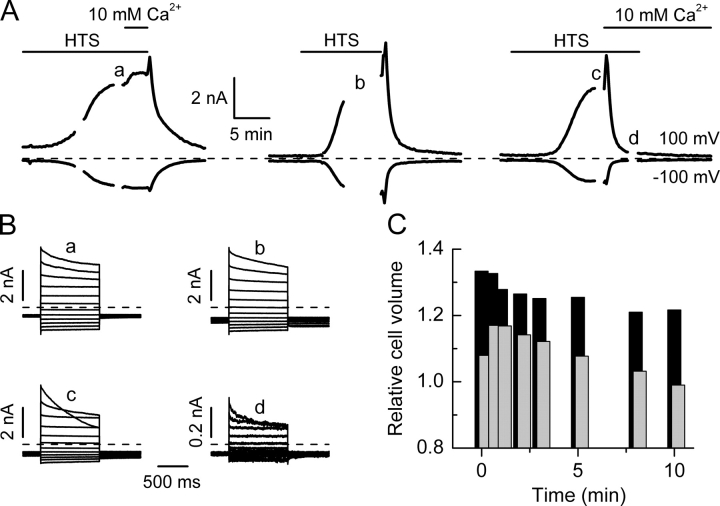
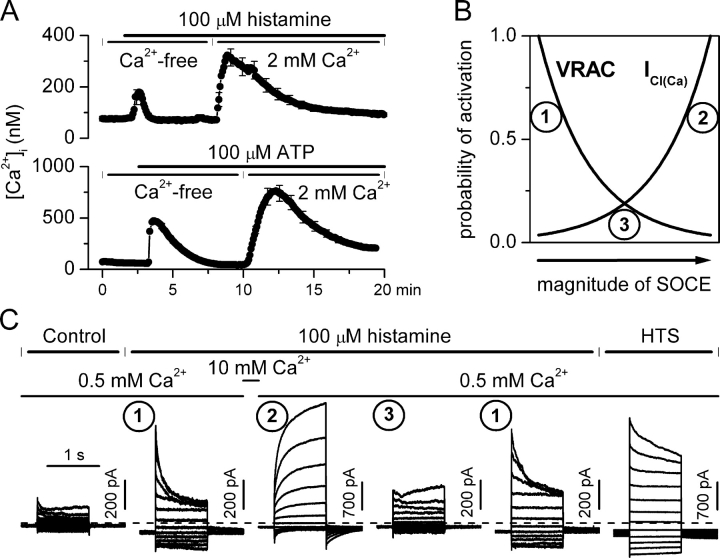
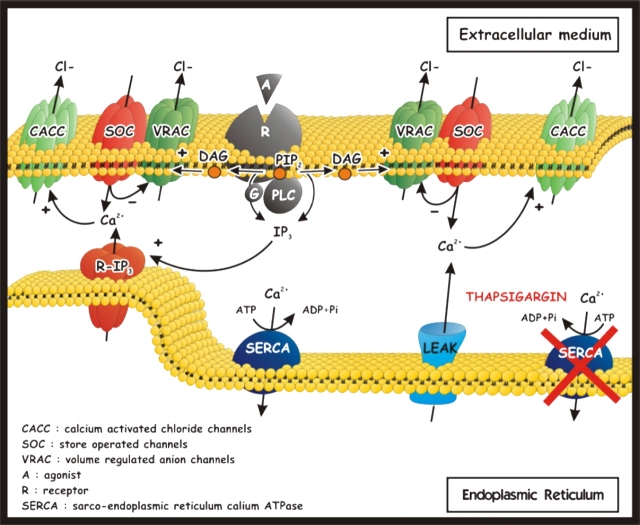
Using patch-clamp and calcium imaging techniques, we characterized the effects of ATP and histamine on human keratinocytes. In the HaCaT cell line, both receptor agonists induced a transient elevation of [Ca2+]i in a Ca(2+)-free medium followed by a secondary [Ca2+]i rise upon Ca2+ readmission due to store-operated calcium entry (SOCE). In voltage-clamped cells, agonists activated two kinetically distinct currents, which showed differing voltage dependences and were identified as Ca(2+)-activated (I(Cl(Ca))) and volume-regulated (I(Cl, swell)) chloride currents. NPPB and DIDS more efficiently inhibited I(Cl(Ca)) and I(Cl, swell), respectively. Cell swelling caused by hypotonic solution invariably activated I(Cl, swell) while regulatory volume decrease occurred in intact cells, as was found in flow cytometry experiments. The PLC inhibitor U-73122 blocked both agonist- and cell swelling-induced I(Cl, swell), while its inactive analogue U-73343 had no effect. I(Cl(Ca)) could be activated by cytoplasmic calcium increase due to thapsigargin (TG)-induced SOCE as well as by buffering [Ca2+]i in the pipette solution at 500 nM. In contrast, I(Cl, swell) could be directly activated by 1-oleoyl-2-acetyl-sn-glycerol (OAG), a cell-permeable DAG analogue, but neither by InsP3 infusion nor by the cytoplasmic calcium increase. PKC also had no role in its regulation. Agonists, OAG, and cell swelling induced I(Cl, swell) in a nonadditive manner, suggesting their convergence on a common pathway. I(Cl, swell) and I(Cl(Ca)) showed only a limited overlap (i.e., simultaneous activation), although various maneuvers were able to induce these currents sequentially in the same cell. TG-induced SOCE strongly potentiated I(Cl(Ca)), but abolished I(Cl, swell), thereby providing a clue for this paradox. Thus, we have established for the first time using a keratinocyte model that I(Cl, swell) can be physiologically activated under isotonic conditions by receptors coupled to the phosphoinositide pathway. These results also suggest a novel function for SOCE, which can operate as a "selection" switch between closely localized channels.
Figures
Similar articles
-
Modulation of aqueous humor outflow by ionic mechanisms involved in trabecular meshwork cell volume regulation.Invest Ophthalmol Vis Sci. 2004 Oct;45(10):3650-61. doi: 10.1167/iovs.04-0060. Invest Ophthalmol Vis Sci. 2004. PMID: 15452073
-
ATP released from astrocytes during swelling activates chloride channels.J Neurophysiol. 2003 Apr;89(4):1870-7. doi: 10.1152/jn.00510.2002. J Neurophysiol. 2003. PMID: 12686569
-
[Adenosine triphosphate-dependence of volume sensitive chloride current in LNCaP cell line of human prostate cancer].Fiziol Zh (1994). 2005;51(1):51-61. Fiziol Zh (1994). 2005. PMID: 15801200 Ukrainian.
-
Role of the Ca2+ -activated Cl- channels bestrophin and anoctamin in epithelial cells.Biol Chem. 2011 Jan;392(1-2):125-34. doi: 10.1515/BC.2011.010. Biol Chem. 2011. PMID: 21194364 Review.
-
Receptor-activated calcium entry channels--who does what, and when?Sci STKE. 2004 Jul 20;2004(243):pe40. doi: 10.1126/stke.2432004pe40. Sci STKE. 2004. PMID: 15280582 Review.
Cited by
-
Anoctamin 1 mediates thermal pain as a heat sensor.Curr Neuropharmacol. 2013 Dec;11(6):641-51. doi: 10.2174/1570159X113119990038. Curr Neuropharmacol. 2013. PMID: 24396339 Free PMC article.
-
Relationship between TMEM16A/anoctamin 1 and LRRC8A.Pflugers Arch. 2016 Oct;468(10):1751-63. doi: 10.1007/s00424-016-1862-1. Epub 2016 Aug 12. Pflugers Arch. 2016. PMID: 27514381
-
The calcium-activated chloride channel anoctamin 1 acts as a heat sensor in nociceptive neurons.Nat Neurosci. 2012 May 27;15(7):1015-21. doi: 10.1038/nn.3111. Nat Neurosci. 2012. PMID: 22634729
-
A FRET sensor of C-terminal movement reveals VRAC activation by plasma membrane DAG signaling rather than ionic strength.Elife. 2019 Jun 18;8:e45421. doi: 10.7554/eLife.45421. Elife. 2019. PMID: 31210638 Free PMC article.
-
Contribution of Anoctamins to Cell Survival and Cell Death.Cancers (Basel). 2019 Mar 19;11(3):382. doi: 10.3390/cancers11030382. Cancers (Basel). 2019. PMID: 30893776 Free PMC article. Review.
References
-
- Abeele, F.V., Y. Shuba, M. Roudbaraki, L. Lemonnier, K. Vanoverberghe, P. Mariot, R. Skryma, and N. Prevarskaya. 2003. Store-operated Ca2+ channels in prostate cancer epithelial cells: function, regulation, and role in carcinogenesis. Cell Calcium. 33:357–373. - PubMed
-
- Biro, T., I. Szabo, L. Kovacs, J. Hunyadi, and L. Csernoch. 1998. Distinct subpopulations in HaCaT cells as revealed by the characteristics of intracellular calcium release induced by phosphoinositide-coupled agonists. Arch. Dermatol. Res. 290:270–276. - PubMed
-
- Boese, S.H., R.K. Kinne, and F. Wehner. 1996. Single-channel properties of swelling-activated anion conductance in rat inner medullary collecting duct cells. Am. J. Physiol. 271:F1224–F1233. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
