

doi: 10.1083/jcb.200409053.
A multistep, GTP-driven mechanism controlling the dynamic cycling of nucleostemin
Affiliations
- PMID: 15657390
- PMCID: PMC2171593
- DOI: 10.1083/jcb.200409053
Item in Clipboard
A multistep, GTP-driven mechanism controlling the dynamic cycling of nucleostemin
J Cell Biol.
.
Abstract
Nucleostemin (NS) was identified as a stem cell- and cancer cell-enriched nucleolar protein that controls the proliferation of these cells. Here, we report the mechanism that regulates its dynamic shuttling between the nucleolus and nucleoplasm. The nucleolar residence of nucleostemin involves a transient and a long-term binding by the basic and GTP-binding domains, and a dissociation mechanism mediated by the COOH-terminal region. This cycle is propelled by the GTP binding state of nucleostemin. We propose that a rapid nucleostemin cycle is designed to translate extra- and intra-cellular signals into the amount of nucleostemin in the nucleolus in a bidirectional and fast manner.
Figures
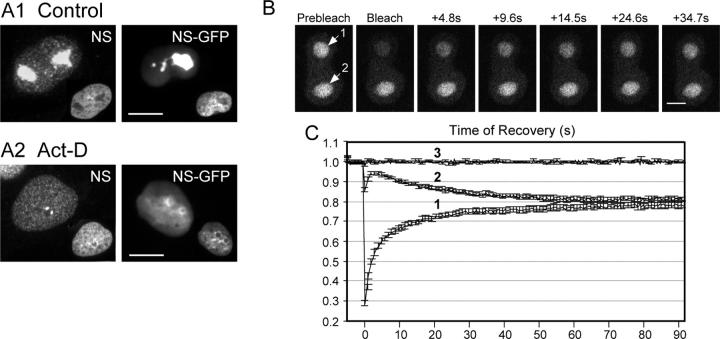
NS cycles between the nucleolus and the nucleoplasm. (A) Endogenous NS and expressed NS-GFP fusion protein display a nucleolar distribution in control cells (A1), and a nucleoplasmic distribution plus intense signals surrounding the nucleolar region in Act-D–treated CHO cells (A2). Inset: DAPI nuclear staining of the same cells. Bars, 10 μm. Time-sequenced images (B) and quantitative analyses (C) of FRAP (label 1) and iFRAP (label 2) on NS-GFP–transfected CHO cells with two nucleoli. After subtraction of the background signal, the relative intensities were normalized to the nonbleached signals in different cells, recorded in parallel, and shown in line 3 (C). Data were averaged over 20 cells in three independent experiments. Y-error bars represent SEM, omitted at every other data point for clarity. Bar, 4 μm.
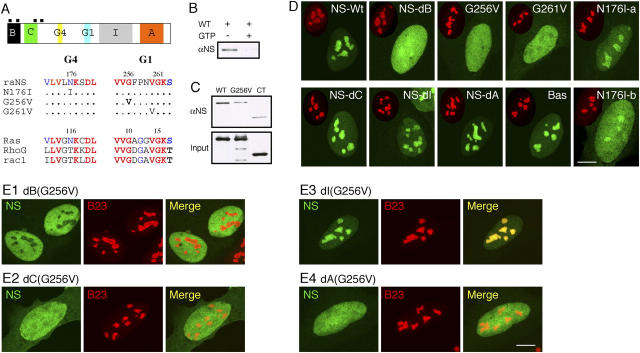
Nucleolar targeting of NS requires the B-domain and GTP-binding domain, and is blocked by the I-domain. (A) Top: a schematic diagram of the NS protein structure (B: basic; C: coiled-coil; G4, G1: GTP-binding motifs; I: intermediate; A: acidic domains; black boxes: nuclear localization signals). (Bottom) Sequences of single amino acid substitution on the conserved residues in the G4 and G1 motifs. (B) Purified GST fusion protein of wild-type NS could bind GTP-agarose (left lane). This interaction was abolished by competition with 10 mM free GTP (right lane). (C) The role of the G1 motif in mediating the GTP binding of NS was shown by a mutation in the G256 residue (G256V) that reduced its binding affinity to the background level as the COOH-terminal NS control (CT). (D) The static distribution of NS mutants in U2 OS cells (NS-Wt: wild-type; NS-dB: B-domain deletion; NS-dC: C-domain deletion; NS-dI: I-domain deletion; NS-dA: A-domain deletion; Bas: B-domain alone). Green: GFP-fused mutants; red: anti-B23 immunostaining of the same cell (scaled to 60%). (E) Mapping the inhibitory domain that gates the nucleolus-targeting event mediated by the B-domain. A deletion of the I-domain (E3), but not the B- (E1), C- (E2), or A- (E4) domain, can rescue the nucleolar phenotype of G256V mutation. Bars (D and E), 10 μm.
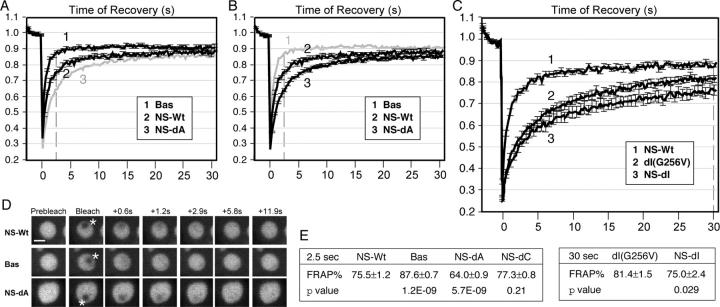
The long-term association of NS in the nucleolus is regulated by the I- and A-domains, and the binding of guanine nucleotide. (A and B) Quantitative data of FRAP analyses of the B-domain alone (Bas, trace 1), the wild-type NS (NS-Wt, trace 2), and the A-domain deletion (NS-dA, trace 3) in CHO cells. Y-error bars represent SEM and are omitted at every other data point for clarity. For comparison, Bas and NS-dA were shown in B and A in gray without error bars. (C) FRAP analyses of dI(G256V) (trace 2) and NS-dI (trace 3), compared with the wild-type NS (trace 1). Dotted lines in A–C indicate the points of analysis. (D) Time-sequenced FRAP images of NS-Wt, Bas, and NS-dA mutants before and after photobleaching. Bar, 4 μm. A 1.5-μm-diameter circle (marked by asterisks) within the nucleolus was bleached and the signal recovery was recorded over a period of 31.5 s. (E) T-test analyses of the FRAP results of wild-type and mutant NS-GFP fusion protein (mean ± SEM, n = 20) were conducted at 2.5 s (left) or 30 s (right) after photobleaching.
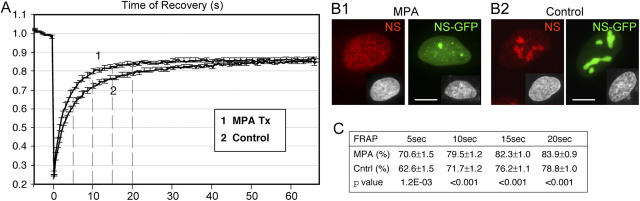
The effect of lowering the intracellular GTP on the static and dynamic distribution of NS. (A) FRAP analyses of NS-GFP in U2 OS cells treated with 40 μM MPA for 24 h (trace 1) or nontreated cells (trace 2). Photobleaching was administered to 3-μm-diameter nucleoli. After subtraction of the background signal, the relative intensities were normalized to the nonbleached signals in different cells, and averaged over 20 cells in three independent experiments. Y-error bars were omitted at every other data points for clarity. Dotted lines indicate the points of analysis. (B) The static distribution of endogenous NS (red), NS-GFP (green), and DAPI nuclear staining (inset) in cells treated with MPA (B1) or untreated cells (B2). Bar, 10 μm. (C) T-tests were conducted at 5, 10, 15, and 20 s after photobleaching to compare between the FRAP rates of MPA-treated cells and the nontreated cells (mean ± SEM).
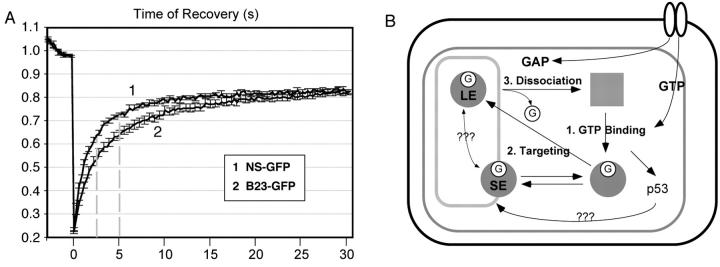
FRAP comparison of NS and B23, and a regulatory model for the dynamic distribution of NS. (A) The nucleolar residence time of NS is shorter than the non-GTP-binding B23. Photobleaching experiments were conducted as described in Fig. 3. Dotted lines indicate the points of analysis. (B) A multistep, GTP-driven model for the regulation of NS distribution. The three rectangular boxes indicate the cell membrane, the nuclear membrane, and the nucleolus (outside-in). The filled circles and square represent the GTP (G)-bound and unbound NS. In the nucleoplasm, the GTP-bound NS is released of the inhibitory activity mediated by the I-domain and can be targeted to the nucleolus. Binding to the nucleolus through the B-domain alone is short lived (SE). The long-term engagement (LE) of NS in the nucleolus involves the GTP-binding domain. Its release requires the I- and A-domains, and may be triggered by the dissociation of guanine nucleotides.
Comment in
-
Going in GTP cycles in the nucleolus.J Cell Biol. 2005 Jan 17;168(2):177-8. doi: 10.1083/jcb.200412038. J Cell Biol. 2005. PMID: 15657389 Free PMC article.
Similar articles
-
Multiple controls regulate nucleostemin partitioning between nucleolus and nucleoplasm.J Cell Sci. 2006 Dec 15;119(Pt 24):5124-36. doi: 10.1242/jcs.03292. J Cell Sci. 2006. PMID: 17158916 Free PMC article.
-
Going in GTP cycles in the nucleolus.J Cell Biol. 2005 Jan 17;168(2):177-8. doi: 10.1083/jcb.200412038. J Cell Biol. 2005. PMID: 15657389 Free PMC article.
-
Depletion of guanine nucleotides leads to the Mdm2-dependent proteasomal degradation of nucleostemin.Cancer Res. 2009 Apr 1;69(7):3004-12. doi: 10.1158/0008-5472.CAN-08-3413. Epub 2009 Mar 24. Cancer Res. 2009. PMID: 19318567 Free PMC article.
-
Nucleolar modulation of TRF1: a dynamic way to regulate telomere and cell cycle by nucleostemin and GNL3L.Cell Cycle. 2009 Sep 15;8(18):2912-6. doi: 10.4161/cc.8.18.9543. Epub 2009 Sep 16. Cell Cycle. 2009. PMID: 19713769 Free PMC article. Review.
-
Nucleophosmin: A Nucleolar Phosphoprotein Orchestrating Cellular Stress Responses.Cells. 2024 Jul 27;13(15):1266. doi: 10.3390/cells13151266. Cells. 2024. PMID: 39120297 Free PMC article. Review.
Cited by
-
Ubiquitin- and MDM2 E3 ligase-independent proteasomal turnover of nucleostemin in response to GTP depletion.J Biol Chem. 2012 Mar 23;287(13):10013-10020. doi: 10.1074/jbc.M111.335141. Epub 2012 Feb 8. J Biol Chem. 2012. PMID: 22318725 Free PMC article.
-
The nucleolus: a model for the organization of nuclear functions.Histochem Cell Biol. 2006 Aug;126(2):135-48. doi: 10.1007/s00418-006-0212-3. Epub 2006 Jul 12. Histochem Cell Biol. 2006. PMID: 16835752 Review.
-
Proteomic Investigation of the Role of Nucleostemin in Nucleophosmin-Mutated OCI-AML 3 Cell Line.Int J Mol Sci. 2022 Jul 11;23(14):7655. doi: 10.3390/ijms23147655. Int J Mol Sci. 2022. PMID: 35886999 Free PMC article.
-
Reactive oxygen species regulate nucleostemin oligomerization and protein degradation.J Biol Chem. 2011 Apr 1;286(13):11035-46. doi: 10.1074/jbc.M110.208470. Epub 2011 Jan 17. J Biol Chem. 2011. PMID: 21242306 Free PMC article.
-
The role of IMP dehydrogenase 2 in Inauhzin-induced ribosomal stress.Elife. 2014 Oct 27;3:e03077. doi: 10.7554/eLife.03077. Elife. 2014. PMID: 25347121 Free PMC article.
References
-
- Blander, G., J. Kipnis, J.F. Leal, C.E. Yu, G.D. Schellenberg, and M. Oren. 1999. Physical and functional interaction between p53 and the Werner's syndrome protein. J. Biol. Chem. 274:29463–29469. - PubMed
-
- Bullough, W.S. 1965. Mitotic and functional homeostasis: a speculative review. Cancer Res. 25:1683–1727. - PubMed
-
- Derenzini, M., D. Trere, A. Pession, M. Govoni, V. Sirri, and P. Chieco. 2000. Nucleolar size indicates the rapidity of cell proliferation in cancer tissues. J. Pathol. 191:181–186. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
