

Spindle-independent condensation-mediated segregation of yeast ribosomal DNA in late anaphase
- PMID: 15657393
- PMCID: PMC2171579
- DOI: 10.1083/jcb.200408087
Spindle-independent condensation-mediated segregation of yeast ribosomal DNA in late anaphase
Abstract
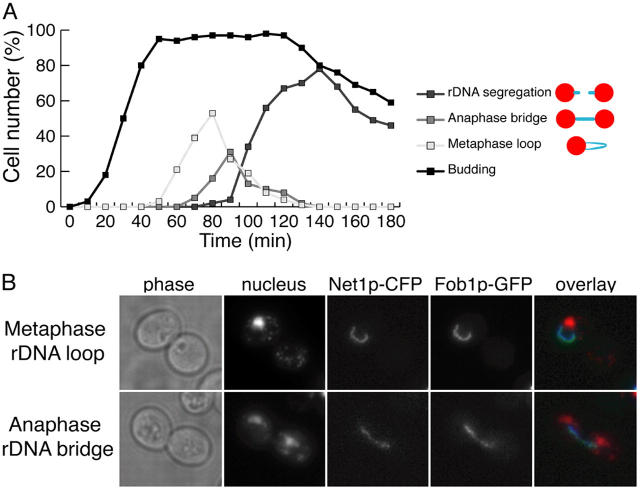
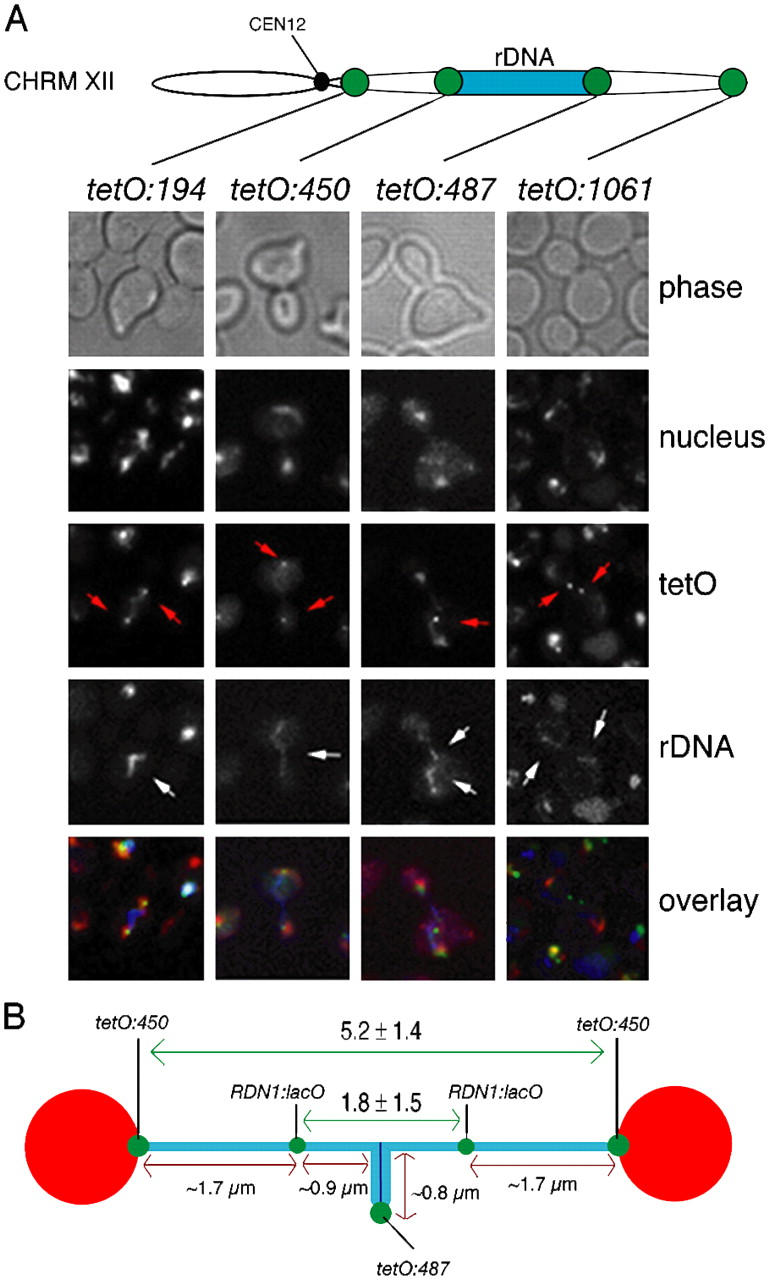
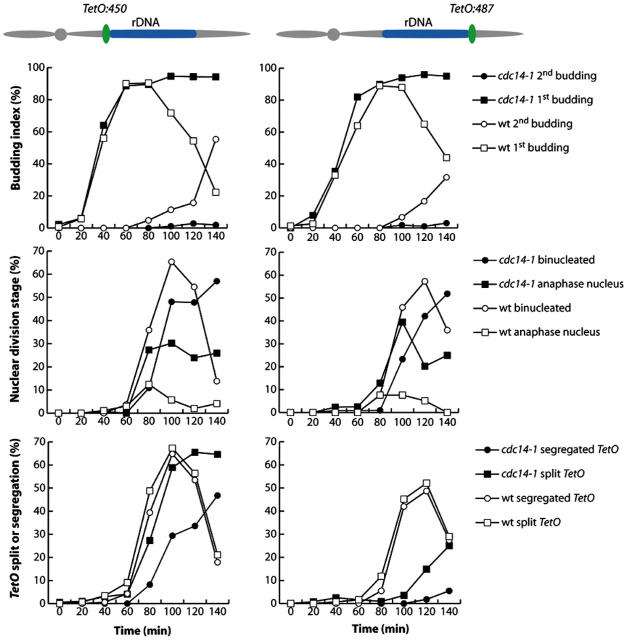
Mitotic cell division involves the equal segregation of all chromosomes during anaphase. The presence of ribosomal DNA (rDNA) repeats on the right arm of chromosome XII makes it the longest in the budding yeast genome. Previously, we identified a stage during yeast anaphase when rDNA is stretched across the mother and daughter cells. Here, we show that resolution of sister rDNAs is achieved by unzipping of the locus from its centromere-proximal to centromere-distal regions. We then demonstrate that during this stretched stage sister rDNA arrays are neither compacted nor segregated despite being largely resolved from each other. Surprisingly, we find that rDNA segregation after this period no longer requires spindles but instead involves Cdc14-dependent rDNA axial compaction. These results demonstrate that chromosome resolution is not simply a consequence of compacting chromosome arms and that overall rDNA compaction is necessary to mediate the segregation of the long arm of chromosome XII.
Figures





Similar articles
-
Cdc14 phosphatase induces rDNA condensation and resolves cohesin-independent cohesion during budding yeast anaphase.Cell. 2004 May 14;117(4):471-82. doi: 10.1016/s0092-8674(04)00415-5. Cell. 2004. PMID: 15137940
-
Cdc14 and the temporal coordination between mitotic exit and chromosome segregation.Cell Cycle. 2005 Jan;4(1):109-12. doi: 10.4161/cc.4.1.1356. Epub 2005 Jan 10. Cell Cycle. 2005. PMID: 15611663 Review.
-
Time course analysis of precocious separation of sister centromeres in budding yeast: continuously separated or frequently reassociated?Genes Cells. 2001 Sep;6(9):765-73. doi: 10.1046/j.1365-2443.2001.00464.x. Genes Cells. 2001. PMID: 11554923
-
Cdc14p/FEAR pathway controls segregation of nucleolus in S. cerevisiae by facilitating condensin targeting to rDNA chromatin in anaphase.Cell Cycle. 2004 Jul;3(7):960-7. doi: 10.4161/cc.3.7.1003. Epub 2004 Jul 4. Cell Cycle. 2004. PMID: 15190202 Free PMC article.
-
Nucleolus in the spotlight.Cell Cycle. 2005 Jan;4(1):106-8. doi: 10.4161/cc.4.1.1355. Epub 2005 Jan 10. Cell Cycle. 2005. PMID: 15611637 Review.
Cited by
-
Altered RNA processing and export lead to retention of mRNAs near transcription sites and nuclear pore complexes or within the nucleolus.Mol Biol Cell. 2016 Sep 1;27(17):2742-56. doi: 10.1091/mbc.E16-04-0244. Epub 2016 Jul 6. Mol Biol Cell. 2016. PMID: 27385342 Free PMC article.
-
Condensin function at centromere chromatin facilitates proper kinetochore tension and ensures correct mitotic segregation of sister chromatids.Genes Cells. 2007 Sep;12(9):1075-90. doi: 10.1111/j.1365-2443.2007.01109.x. Genes Cells. 2007. PMID: 17825050 Free PMC article.
-
Analysis of rRNA Gene Methylation in Arabidopsis thaliana by CHEF-Conventional 2D Gel Electrophoresis.Methods Mol Biol. 2016;1455:183-202. doi: 10.1007/978-1-4939-3792-9_14. Methods Mol Biol. 2016. PMID: 27576719 Free PMC article.
-
Focal accumulation of preribosomes outside the nucleolus during metaphase-anaphase in budding yeast.RNA. 2017 Sep;23(9):1432-1443. doi: 10.1261/rna.061259.117. Epub 2017 Jun 6. RNA. 2017. PMID: 28588079 Free PMC article.
-
Pds5 regulators segregate cohesion and condensation pathways in Saccharomyces cerevisiae.Proc Natl Acad Sci U S A. 2015 Jun 2;112(22):7021-6. doi: 10.1073/pnas.1501369112. Epub 2015 May 18. Proc Natl Acad Sci U S A. 2015. PMID: 25986377 Free PMC article.
References
-
- Alexandru, G., F. Uhlmann, K. Mechtler, M.A. Poupart, and K. Nasmyth. 2001. Phosphorylation of the cohesin subunit Scc1 by Polo/Cdc5 kinase regulates sister chromatid separation in yeast. Cell. 105:459–472. - PubMed
-
- Buonomo, S.B., K.P. Rabitsch, J. Fuchs, S. Gruber, M. Sullivan, F. Uhlmann, M. Petronczki, A. Toth, and K. Nasmyth. 2003. Division of the nucleolus and its release of CDC14 during anaphase of meiosis I depends on separase, SPO12, and SLK19. Dev. Cell. 4:727–739. - PubMed
-
- D'Amours, D., F. Stegmeier, and A. Amon. 2004. Cdc14 and condensin control the dissolution of cohesin-independent chromosome linkages at repeated DNA. Cell. 117:455–469. - PubMed
-
- Darlington, C.D. 1937. Recent Advances in Cytology. J&A Churchill Ltd., London. 356 pp.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
