

Similar effects of Brca2 truncation and Rad51 paralog deficiency on immunoglobulin V gene diversification in DT40 cells support an early role for Rad51 paralogs in homologous recombination
- PMID: 15657438
- PMCID: PMC544004
- DOI: 10.1128/MCB.25.3.1124-1134.2005
Similar effects of Brca2 truncation and Rad51 paralog deficiency on immunoglobulin V gene diversification in DT40 cells support an early role for Rad51 paralogs in homologous recombination
Abstract
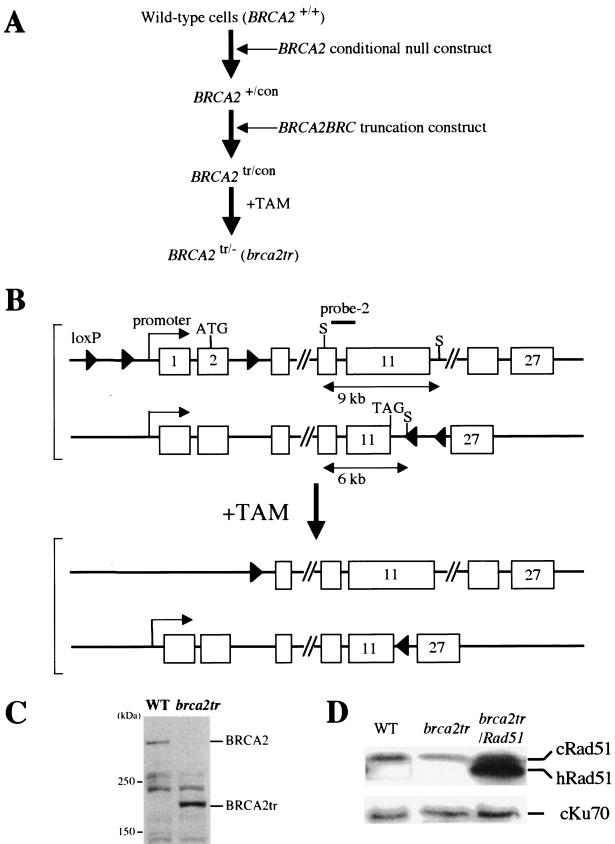
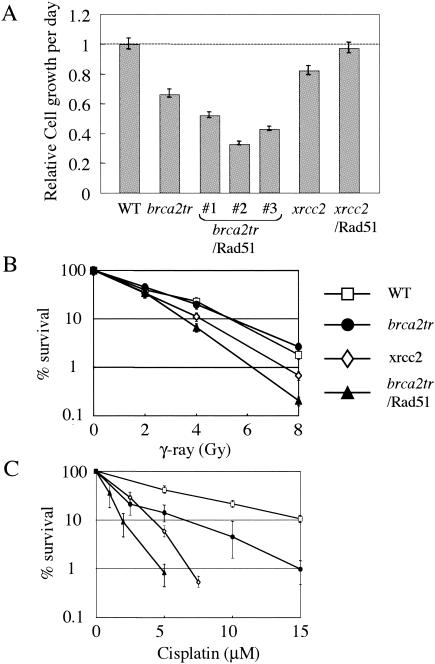
BRCA2 is a tumor suppressor gene that is linked to hereditary breast and ovarian cancer. Although the Brca2 protein participates in homologous DNA recombination (HR), its precise role remains unclear. From chicken DT40 cells, we generated BRCA2 gene-deficient cells which harbor a truncation at the 3' end of the BRC3 repeat (brca2tr). Comparison of the characteristics of brca2tr cells with those of other HR-deficient DT40 clones revealed marked similarities with rad51 paralog mutants (rad51b, rad51c, rad51d, xrcc2, or xrcc3 cells). The phenotypic similarities include a shift from HR-mediated diversification to single-nucleotide substitutions in the immunoglobulin variable gene segment and the partial reversion of this shift by overexpression of Rad51. Although recent evidence supports at least Xrcc3 and Rad51C playing a role late in HR, our data suggest that Brca2 and the Rad51 paralogs may also contribute to HR at the same early step, with their loss resulting in the stimulation of an alternative, error-prone repair pathway.
Figures




Similar articles
-
Rad51 paralog complexes BCDX2 and CX3 act at different stages in the BRCA1-BRCA2-dependent homologous recombination pathway.Mol Cell Biol. 2013 Jan;33(2):387-95. doi: 10.1128/MCB.00465-12. Epub 2012 Nov 12. Mol Cell Biol. 2013. PMID: 23149936 Free PMC article.
-
Homologous recombination-deficient mutation cluster in tumor suppressor RAD51C identified by comprehensive analysis of cancer variants.Proc Natl Acad Sci U S A. 2022 Sep 20;119(38):e2202727119. doi: 10.1073/pnas.2202727119. Epub 2022 Sep 13. Proc Natl Acad Sci U S A. 2022. PMID: 36099300 Free PMC article.
-
RAD51 paralogs promote homology-directed repair at diversifying immunoglobulin V regions.BMC Mol Biol. 2009 Oct 28;10:98. doi: 10.1186/1471-2199-10-98. BMC Mol Biol. 2009. PMID: 19863810 Free PMC article.
-
RAD51 paralogs: roles in DNA damage signalling, recombinational repair and tumorigenesis.Semin Cell Dev Biol. 2011 Oct;22(8):898-905. doi: 10.1016/j.semcdb.2011.07.019. Epub 2011 Jul 28. Semin Cell Dev Biol. 2011. PMID: 21821141 Review.
-
RAD-ical New Insights into RAD51 Regulation.Genes (Basel). 2018 Dec 13;9(12):629. doi: 10.3390/genes9120629. Genes (Basel). 2018. PMID: 30551670 Free PMC article. Review.
Cited by
-
Genetic variation stimulated by epigenetic modification.PLoS One. 2008;3(12):e4075. doi: 10.1371/journal.pone.0004075. Epub 2008 Dec 30. PLoS One. 2008. PMID: 19115012 Free PMC article.
-
Genetic evidence for single-strand lesions initiating Nbs1-dependent homologous recombination in diversification of Ig v in chicken B lymphocytes.PLoS Genet. 2009 Jan;5(1):e1000356. doi: 10.1371/journal.pgen.1000356. Epub 2009 Jan 30. PLoS Genet. 2009. PMID: 19180185 Free PMC article.
-
Brca1 in immunoglobulin gene conversion and somatic hypermutation.DNA Repair (Amst). 2008 Feb 1;7(2):253-66. doi: 10.1016/j.dnarep.2007.10.002. Epub 2007 Nov 26. DNA Repair (Amst). 2008. PMID: 18036997 Free PMC article.
-
Trapping of PARP1 and PARP2 by Clinical PARP Inhibitors.Cancer Res. 2012 Nov 1;72(21):5588-99. doi: 10.1158/0008-5472.CAN-12-2753. Cancer Res. 2012. PMID: 23118055 Free PMC article.
-
MSH6- or PMS2-deficiency causes re-replication in DT40 B cells, but it has little effect on immunoglobulin gene conversion or on repair of AID-generated uracils.Nucleic Acids Res. 2013 Mar 1;41(5):3032-46. doi: 10.1093/nar/gks1470. Epub 2013 Jan 11. Nucleic Acids Res. 2013. PMID: 23314153 Free PMC article.
References
-
- Abbott, D. W., M. L. Freeman, and J. T. Holt. 1998. Double-strand break repair deficiency and radiation sensitivity in BRCA2 mutant cancer cells. J. Natl. Cancer Inst. 90:978-985. - PubMed
-
- Arakawa, H., J. Hauschild, and J. M. Buerstedde. 2002. Requirement of the activation-induced deaminase (AID) gene for immunoglobulin gene conversion. Science 295:1301-1306. - PubMed
-
- Baumann, P., F. E. Benson, and S. C. West. 1996. Human Rad51 protein promotes ATP-dependent homologous pairing and strand transfer reactions in vitro. Cell 87:757-766. - PubMed
-
- Bezzubova, O., A. Silbergleit, Y. Yamaguchi-Iwai, S. Takeda, and J. M. Buerstedde. 1997. Reduced X-ray resistance and homologous recombination frequencies in a RAD54−/− mutant of the chicken DT40 cell line. Cell 89:185-193. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous