

The snRNP 15.5K protein folds its cognate K-turn RNA: a combined theoretical and biochemical study
- PMID: 15659359
- PMCID: PMC1370708
- DOI: 10.1261/rna.7149605
The snRNP 15.5K protein folds its cognate K-turn RNA: a combined theoretical and biochemical study
Abstract
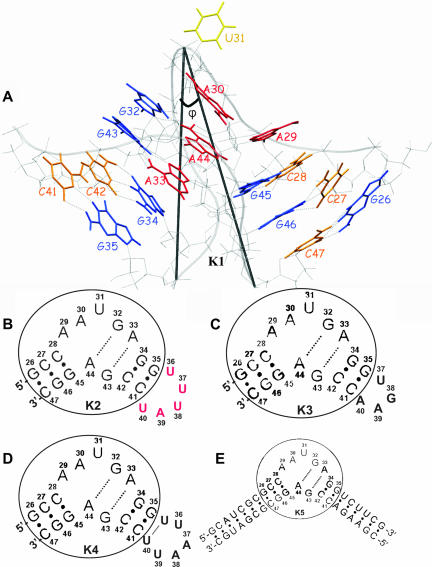
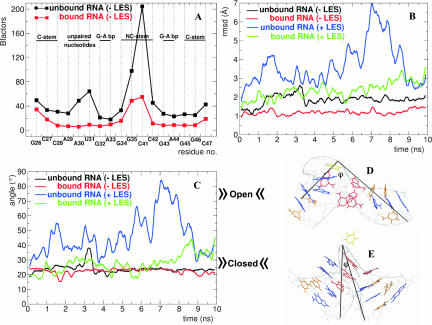
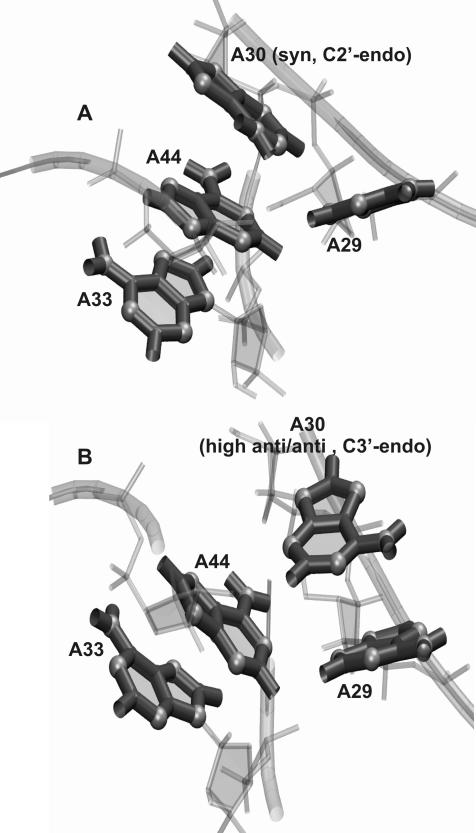
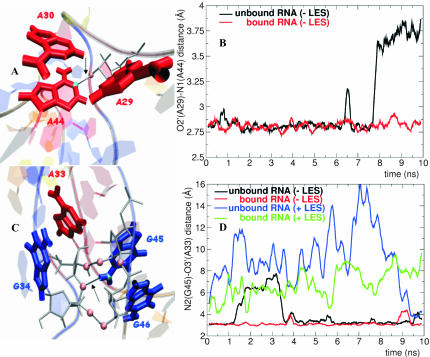
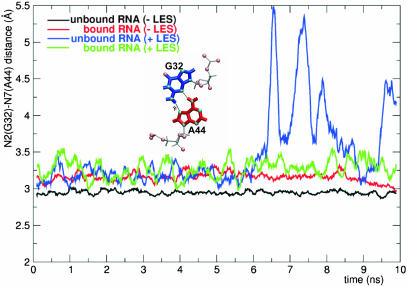
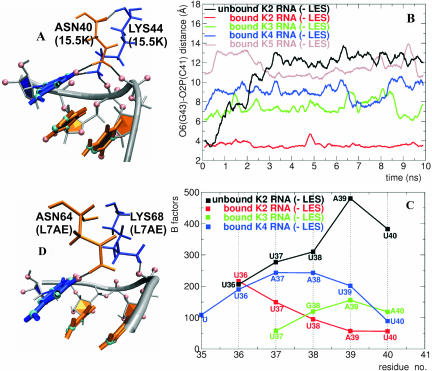
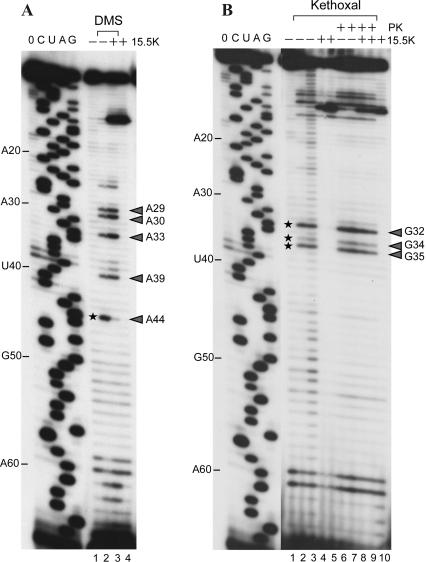
The human 15.5K protein binds to the 5' stem-loop of U4 snRNA, promotes the assembly of the spliceosomal U4/U6 snRNP, and is required for the recruitment of the 61K protein and the 20/60/90K protein complex to the U4 snRNA. In the crystallographic structure of the 15.5K-U4 snRNA complex, the conformation of the RNA corresponds to the family of kink-turn (K-turn) structural motifs. We simulated the complex and the free RNA, showing how the protein binding and the intrinsic flexibility contribute to the RNA folding process. We found that the RNA is significantly more flexible in the absence of the 15.5K protein. Conformational transitions such as the interconversion between alternative purine stacking schemes, the loss of G-A base pairs, and the opening of the K-turn occur only in the free RNA. Furthermore, the stability of one canonical G-C base pair is influenced both by the binding of the 15.5K protein and the nature of the adjacent structural element in the RNA. We performed chemical RNA modification experiments and observed that the free RNA lacks secondary structure elements, a result in excellent agreement with the simulations. Based on these observations, we propose a protein-assisted RNA folding mechanism in which the RNA intrinsic flexibility functions as a catalyst.
Figures
Similar articles
-
Hierarchical, clustered protein interactions with U4/U6 snRNA: a biochemical role for U4/U6 proteins.EMBO J. 2002 Oct 15;21(20):5527-38. doi: 10.1093/emboj/cdf544. EMBO J. 2002. PMID: 12374753 Free PMC article.
-
Structural basis for the dual U4 and U4atac snRNA-binding specificity of spliceosomal protein hPrp31.RNA. 2011 Sep;17(9):1655-63. doi: 10.1261/rna.2690611. Epub 2011 Jul 22. RNA. 2011. PMID: 21784869 Free PMC article.
-
RNA structural requirements for the association of the spliceosomal hPrp31 protein with the U4 and U4atac small nuclear ribonucleoproteins.J Biol Chem. 2006 Sep 22;281(38):28278-86. doi: 10.1074/jbc.M603350200. Epub 2006 Jul 20. J Biol Chem. 2006. PMID: 16857676
-
Spliceosome assembly in the absence of stable U4/U6 RNA pairing.RNA. 2015 May;21(5):923-34. doi: 10.1261/rna.048421.114. Epub 2015 Mar 11. RNA. 2015. PMID: 25762536 Free PMC article.
-
CryoEM structures of two spliceosomal complexes: starter and dessert at the spliceosome feast.Curr Opin Struct Biol. 2016 Feb;36:48-57. doi: 10.1016/j.sbi.2015.12.005. Epub 2016 Jan 21. Curr Opin Struct Biol. 2016. PMID: 26803803 Free PMC article. Review.
Cited by
-
Elastic properties of ribosomal RNA building blocks: molecular dynamics of the GTPase-associated center rRNA.Nucleic Acids Res. 2007;35(12):4007-17. doi: 10.1093/nar/gkm245. Epub 2007 Jun 6. Nucleic Acids Res. 2007. PMID: 17553840 Free PMC article.
-
Recurrent structural RNA motifs, Isostericity Matrices and sequence alignments.Nucleic Acids Res. 2005 Apr 28;33(8):2395-409. doi: 10.1093/nar/gki535. Print 2005. Nucleic Acids Res. 2005. PMID: 15860776 Free PMC article.
-
Conformational dynamics of RNA-peptide binding: a molecular dynamics simulation study.Biophys J. 2006 Jan 15;90(2):391-9. doi: 10.1529/biophysj.105.069559. Epub 2005 Oct 20. Biophys J. 2006. PMID: 16239331 Free PMC article.
-
Dynamic guide-target interactions contribute to sequential 2'-O-methylation by a unique archaeal dual guide box C/D sRNP.RNA. 2008 Jul;14(7):1411-23. doi: 10.1261/rna.1003308. Epub 2008 May 30. RNA. 2008. PMID: 18515549 Free PMC article.
-
Biochemical defects in minor spliceosome function in the developmental disorder MOPD I.RNA. 2014 Jul;20(7):1078-89. doi: 10.1261/rna.045187.114. Epub 2014 May 27. RNA. 2014. PMID: 24865609 Free PMC article.
References
-
- Aboulela, F. and Varani, G. 1998. Recognition of HIV-1 TAR RNA by Tat protein and Tat-derived peptides. J. Mol. Struct. 423: 29–39.
-
- Auffinger, P. and Westhof, E. 2000. Water and ion binding around RNA and DNA (C,G) oligomers. J. Mol. Biol. 300: 1113–1131. - PubMed
-
- Auffinger, P., Bielecki, L., and Westhof, E. 2003. The Mg2+ binding sites of the 5S rRNA loop E motif as investigated by molecular dynamics simulations. Chem. Biol. 10: 551–561. - PubMed
-
- Auffinger, P., Bielecki, L., and Westhof, E. 2004a. Symmetric K+ and Mg2+ ionbinding sites in the 5S rRNA loop E inferred from molecular dynamics simulations. J. Mol. Biol. 335: 555–571. - PubMed
-
- ———. 2004b. Anion binding to nucleic acids. Structure 12: 379–388. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous