

Acetic acid induces expression of the Staphylococcus aureus cidABC and lrgAB murein hydrolase regulator operons
- PMID: 15659658
- PMCID: PMC545714
- DOI: 10.1128/JB.187.3.813-821.2005
Acetic acid induces expression of the Staphylococcus aureus cidABC and lrgAB murein hydrolase regulator operons
Abstract
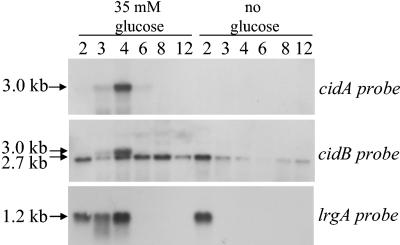
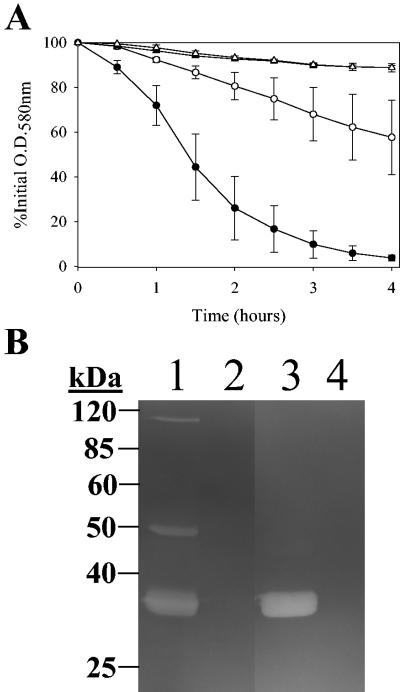
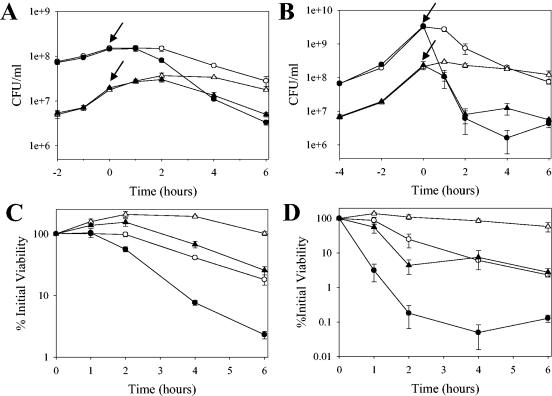
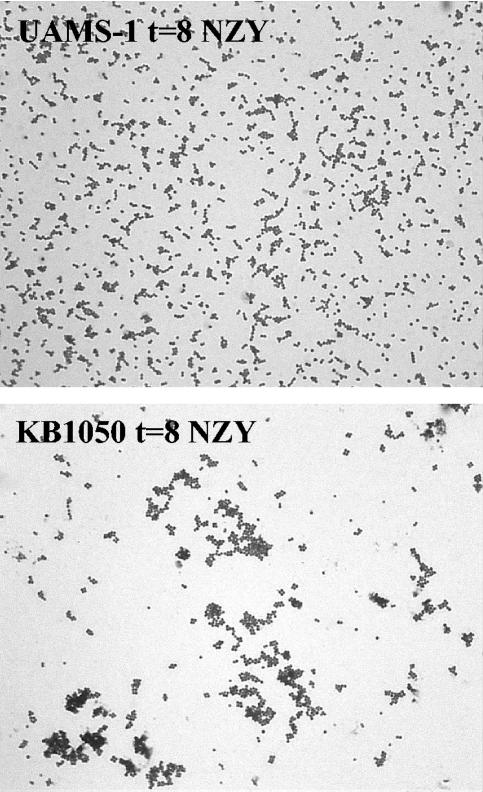
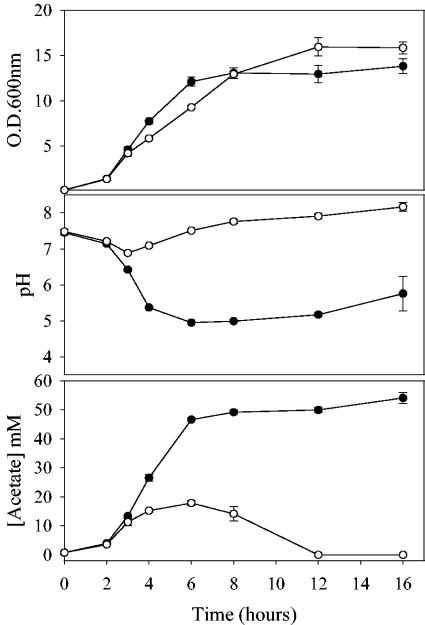
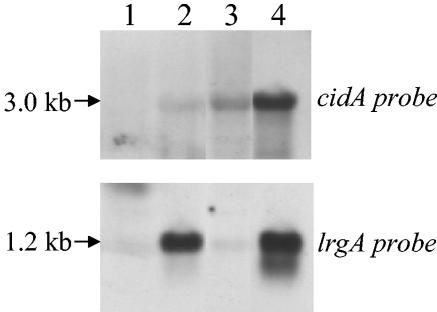
The Staphylococcus aureus lrg and cid operons encode homologous proteins that regulate extracellular murein hydrolase activity and penicillin tolerance in a diametrically opposing manner. Although their specific regulatory functions remain unknown, it has been postulated that the functions of CidA and LrgA are analogous to those of bacteriophage holins and antiholins, respectively, and that these proteins serve as molecular control elements of bacterial programmed cell death. Although these studies demonstrated that cidBC transcription is abundant in sigmaB-proficient strains, cidABC transcription was only minimally expressed under standard growth conditions. In this study, we demonstrate that cidABC and lrgAB transcription in the clinical isolate UAMS-1 is induced by growth in the presence of 35 mM glucose and that this enhances murein hydrolase activity and decreases tolerance to vancomycin and rifampin. The effect of glucose on murein hydrolase activity was not observed in the cidA mutant, indicating that the induction of this activity was dependent on enhanced cidABC expression. Furthermore, we demonstrate that the effects of glucose on cidABC and lrgAB transcription are mediated by the generation of acetic acid produced by the metabolism of this and other carbon sources. These results shed new light on the control of the S. aureus cidABC and lrgAB genes and demonstrate that these operons, as well as murein hydrolase activity and antibiotic tolerance, are responsive to carbohydrate metabolism.
Figures

References
-
- Abdel-Hamid, A. M., M. M. Attwood, and J. R. Guest. 2001. Pyruvate oxidase contributes to the aerobic growth efficiency of Escherichia coli. Microbiology 147:1483-1498. - PubMed
-
- Axe, D. D., and J. E. Bailey. 1995. Transport of lactate and acetate through the energized cytoplasmic membrane of Escherichia coli. Biotechnol. Bioeng. 47:8-19. - PubMed
-
- Bayles, K. W. 2003. Are the molecular strategies that control apoptosis conserved in bacteria? Trends Microbiol. 11:306-311. - PubMed
-
- Bayles, K. W. 2000. The bactericidal action of penicillin: new clues to an unsolved mystery. Trends Microbiol. 8:274-278. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
