

Relative contribution of chloride channels and transporters to regulatory volume decrease in human glioma cells
- PMID: 15659714
- PMCID: PMC2548409
- DOI: 10.1152/ajpcell.00503.2004
Relative contribution of chloride channels and transporters to regulatory volume decrease in human glioma cells
Abstract
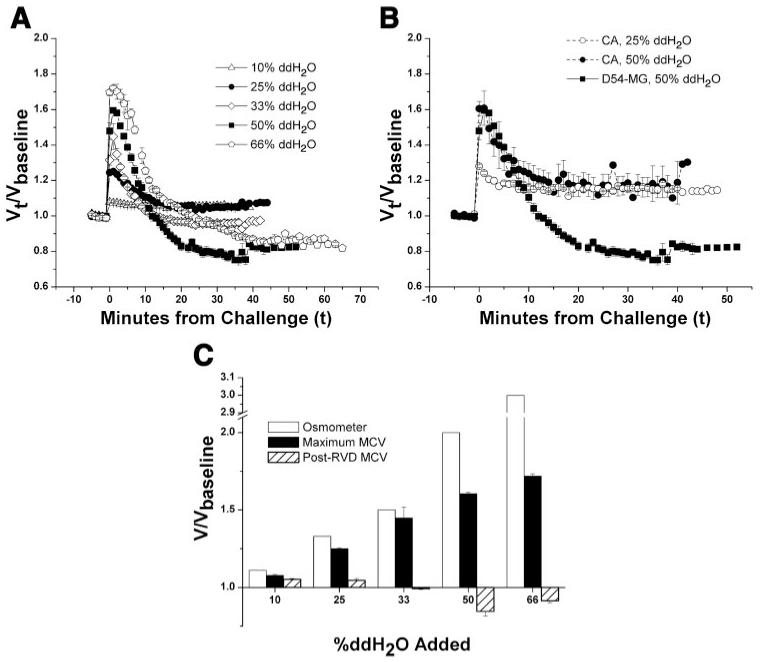
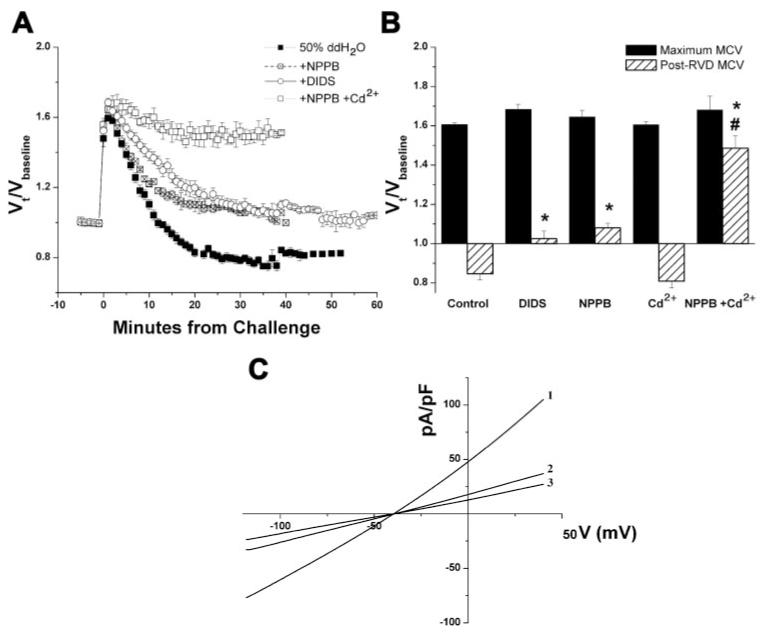
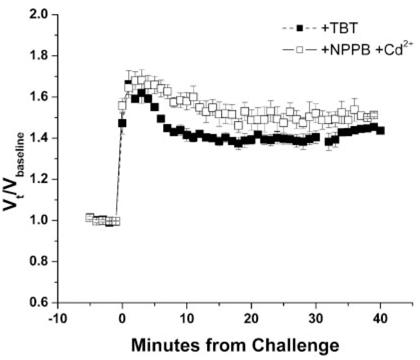
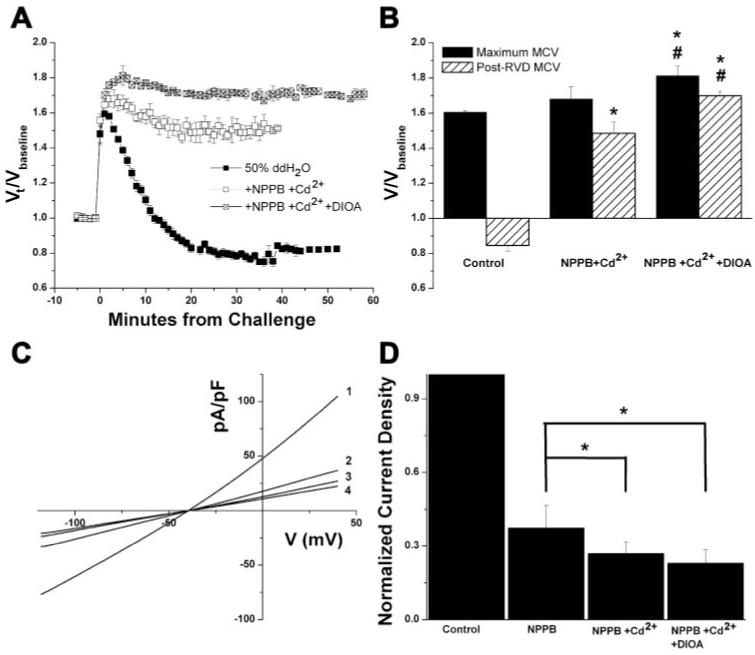
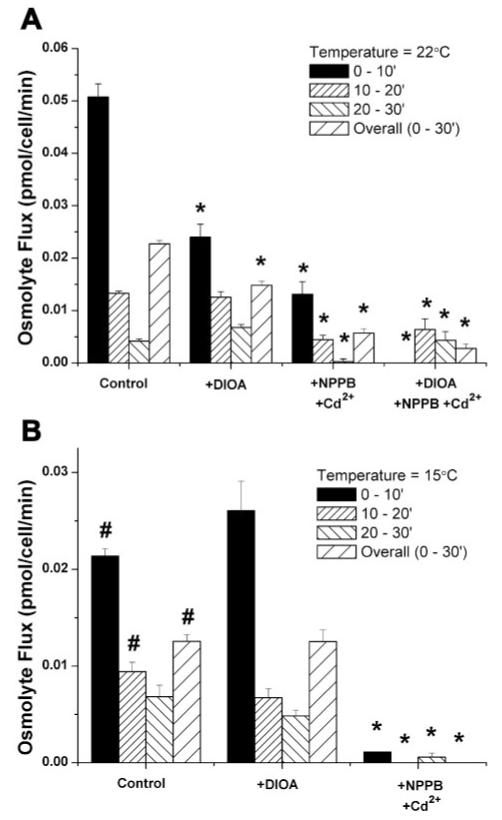
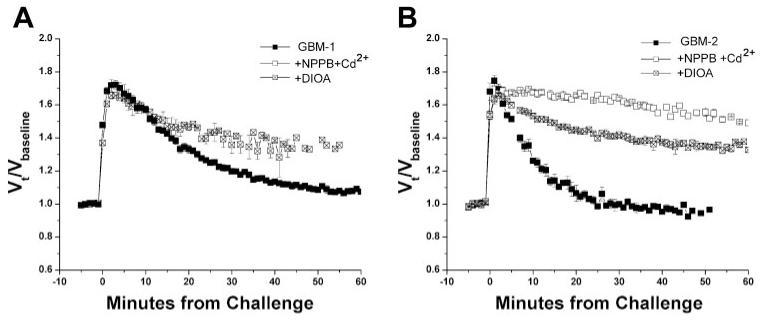
Primary brain tumors (gliomas) often present with peritumoral edema. Their ability to thrive in this osmotically altered environment prompted us to examine volume regulation in human glioma cells, specifically the relative contribution of Cl(-) channels and transporters to this process. After a hyposmotic challenge, cultured astrocytes, D54-MG glioma cells, and glioma cells from human patient biopsies exhibited a regulatory volume decrease (RVD). Although astrocytes were not able to completely reestablish their original prechallenge volumes, glioma cells exhibited complete volume recovery, sometimes recovering to a volume smaller than their original volumes (V(Post-RVD) < V(baseline)). In glioma cells, RVD was largely inhibited by treatment with a combination of Cl(-) channel inhibitors, 5-nitro-2-(3-phenylpropylamino)benzoic acid (NPPB) and Cd(2+) (V(Post-RVD) > 1.4*V(baseline)). Volume regulation was also attenuated to a lesser degree by the addition of R-(+)-[(2-n-butyl-6,7-dichloro-2-cyclopentyl-2,3-dihydro-1-oxo-1H-inden-5-yl)oxy]acetic acid (DIOA), a known K(+)-Cl(-) cotransporter (KCC) inhibitor. To dissect the relative contribution of channels vs. transporters in RVD, we took advantage of the comparatively high temperature dependence of transport processes vs. channel-mediated diffusion. Cooling D54-MG glioma cells to 15 degrees C resulted in a loss of DIOA-sensitive volume regulation. Moreover, at 15 degrees C, the channel blockers NPPB + Cd(2+) completely inhibited RVD and cells behaved like perfect osmometers. The calculated osmolyte flux during RVD under these experimental conditions suggests that the relative contribution of Cl(-) channels vs. transporters to this process is approximately 60-70% and approximately 30-40%, respectively. Finally, we identified several candidate proteins that may be involved in RVD, including the Cl(-) channels ClC-2, ClC-3, ClC-5, ClC-6, and ClC-7 and the transporters KCC1 and KCC3a.
Figures



References
-
- Botchkin LM, Matthews G. Swelling activates chloride current and increases internal calcium in nonpigmented epithelial cells from the rabbit ciliary body. J Cell Physiol. 1995;164:286–294. - PubMed
-
- Brauer M, Frei E, Claes L, Grissmer S, Jager H. Influence of K-Cl cotransporter activity on activation of volume-sensitive Cl- channels in human osteoblasts. Am J Physiol Cell Physiol. 2003;285:C22–C30. - PubMed
-
- de Tassigny A D’Anglemont, Souktani R, Ghaleh B, Henry P, Berdeaux A. Structure and pharmacology of swelling-sensitive chloride channels, ICl,swell. Fundam Clin Pharmacol. 2003;17:539–553. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical