

Genetics and geometry of canalization and developmental stability in Drosophila subobscura
- PMID: 15663797
- PMCID: PMC548280
- DOI: 10.1186/1471-2148-5-7
Genetics and geometry of canalization and developmental stability in Drosophila subobscura
Abstract
Background: Many properties of organisms show great robustness against genetic and environmental perturbations. The terms canalization and developmental stability were originally proposed to describe the ability of an organism to resist perturbations and to produce a predictable target phenotype regardless of random developmental noise. However, the extent to which canalization and developmental stability are controlled by the same set of genes and share underlying regulatory mechanisms is largely unresolved.
Results: We have analyzed the effects of clinical genetic variation (inversion polymorphism) on wing asymmetry by applying the methods of geometric morphometrics in the context of quantitative genetics using isochromosomal lines of Drosophila subobscura. For the analysis of overall size, developmental stability was positively correlated with levels of heterozygosity and development at the optimal temperature. For analyses of shape, the overall comparisons by matrix correlations indicate that inter- and intraindividual variation levels were poorly correlated, a result also supported when comparing the vectors describing patterns of variation of landmark position. The lack of similarity was basically due to the discrepancy between the genetic and environmental components of the interindividual variation. Finally, the analyses have also underscored the presence of genetic variation for directional asymmetry.
Conclusions: The results strongly support the hypothesis that environmental canalization and developmental stability share underlying regulatory mechanisms, but environmental and genetic canalization are not functionally the same. A likely explanation for this lack of association is that natural wing shape variation in Drosophila populations is loosely related to individual fitness.
Figures
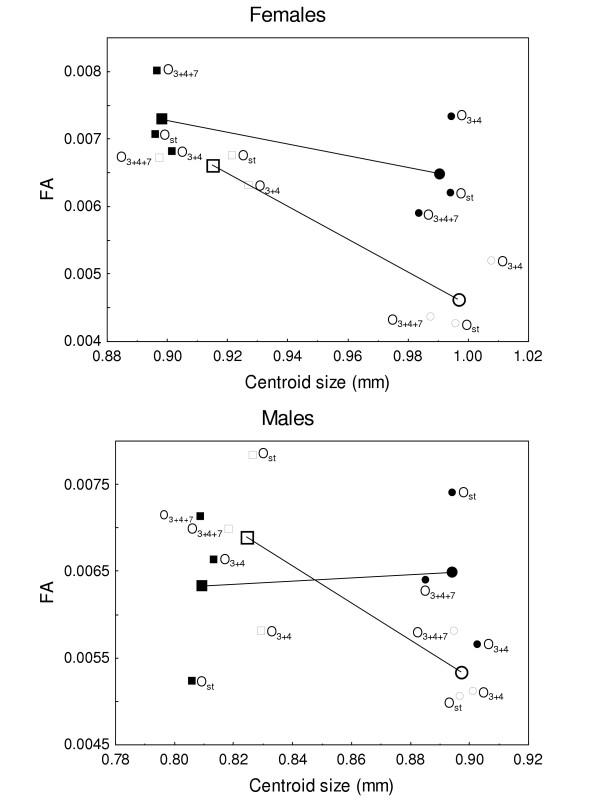
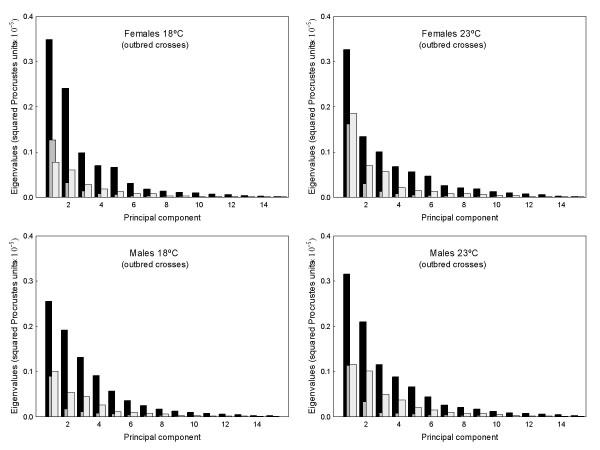
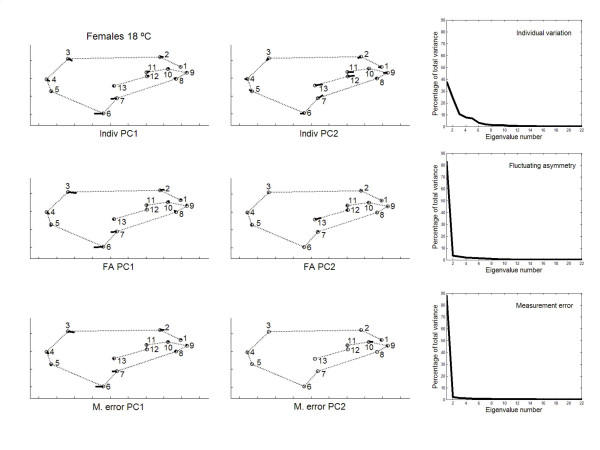
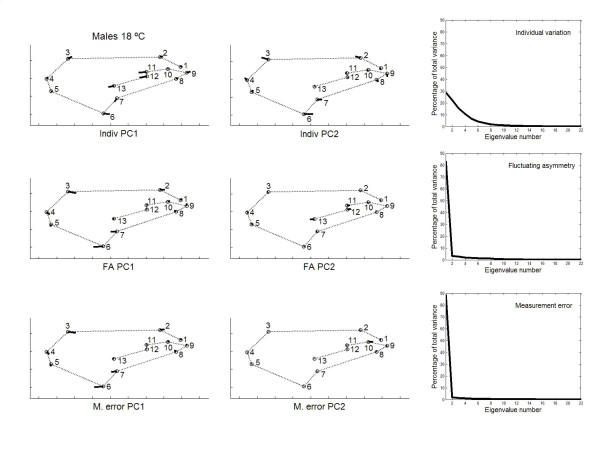
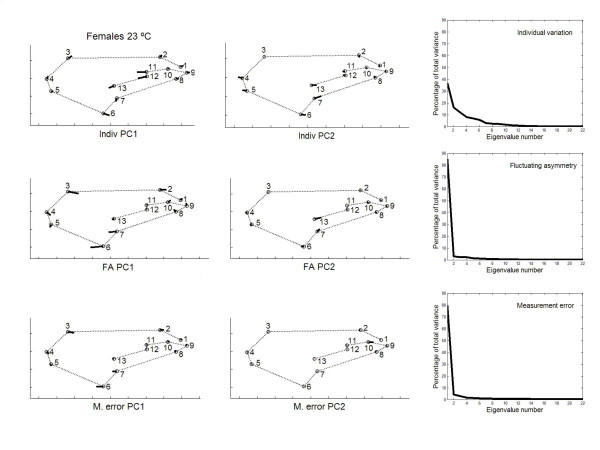
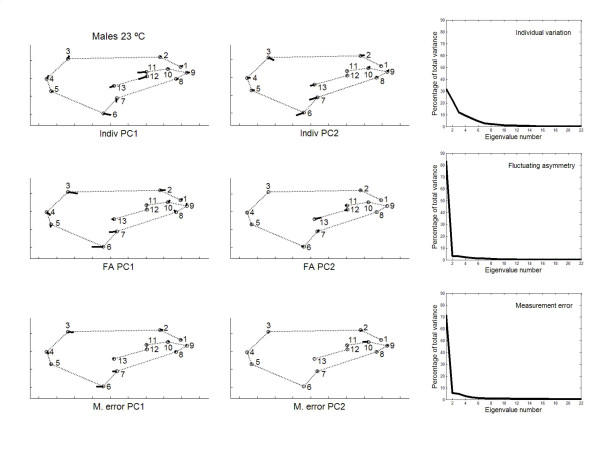
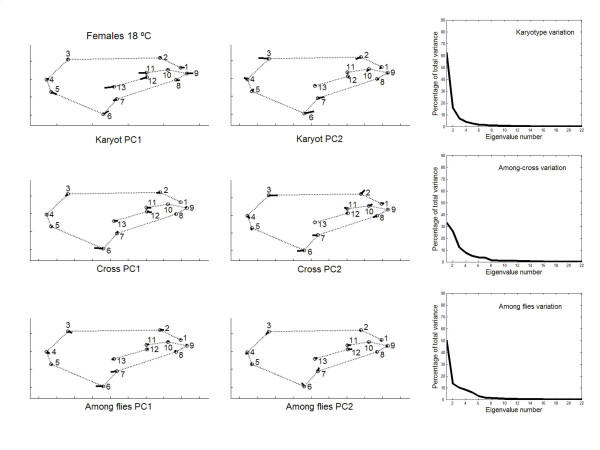
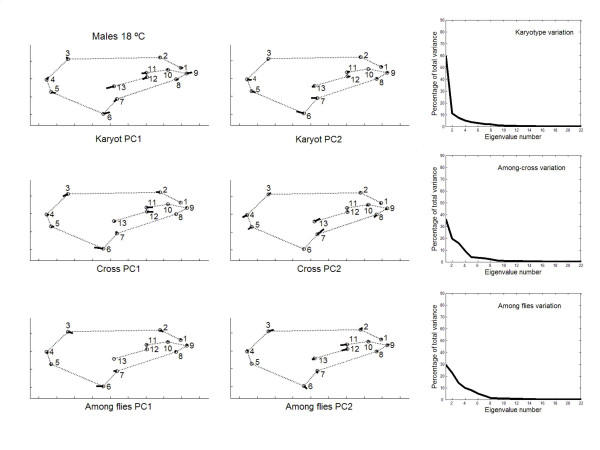
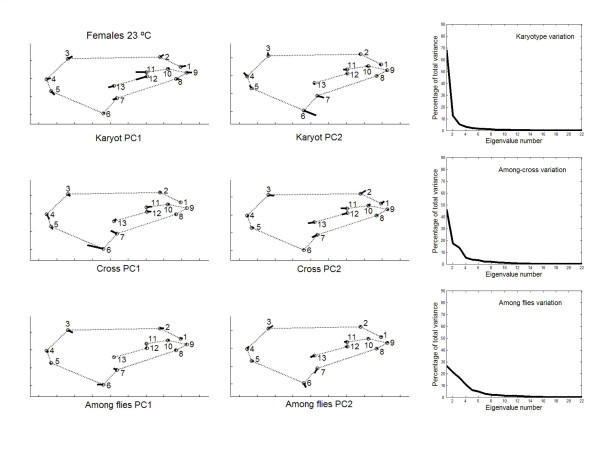
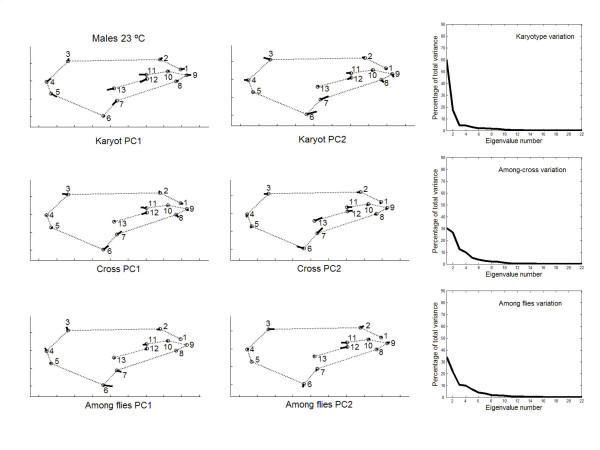
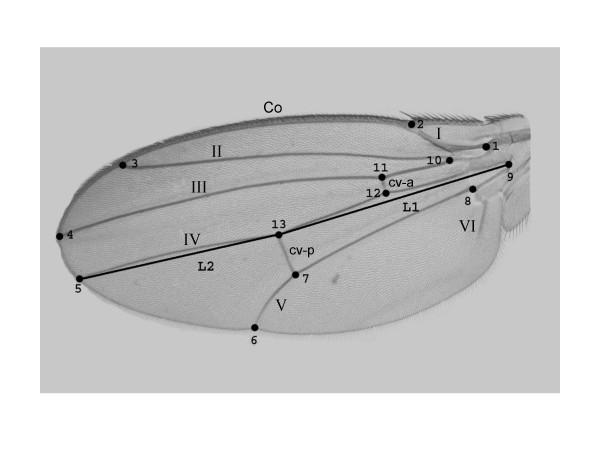
 has been previously used to study shape clines in this species [30].
has been previously used to study shape clines in this species [30].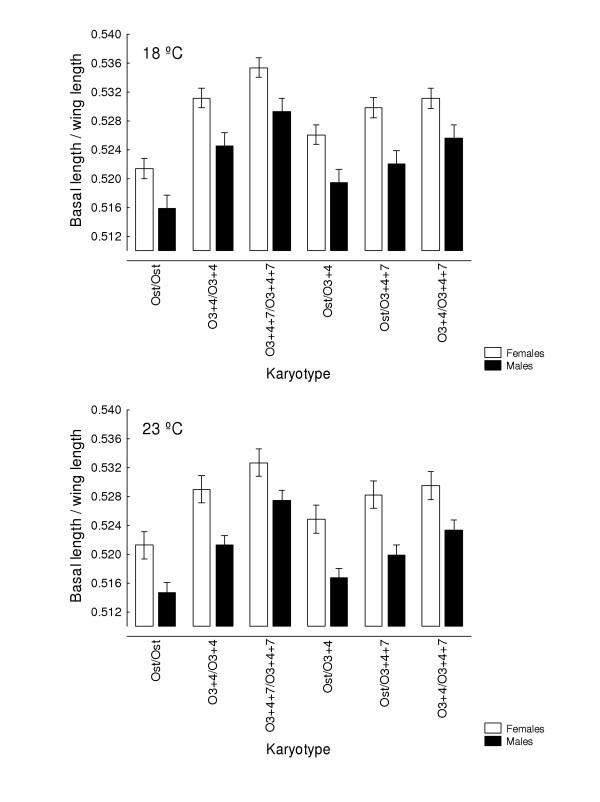
 , with karyotype and temperature as fixed effects, and crosses nested within karyotypes, detected statistically significant differences for the main effects (karyotype: female F5,30 = 12.625, P < 0.001; male F5,30 = 9.785, P < 0.001. Temperature: female F1,390 = 30.219, P < 0.001; male F1,390 = 61.835, P < 0.001) but no karyotype × temperature interaction (females: F5,390 = 1.570, P = 0.168; males: F 5,390 = 1.111, P = 0.354).
, with karyotype and temperature as fixed effects, and crosses nested within karyotypes, detected statistically significant differences for the main effects (karyotype: female F5,30 = 12.625, P < 0.001; male F5,30 = 9.785, P < 0.001. Temperature: female F1,390 = 30.219, P < 0.001; male F1,390 = 61.835, P < 0.001) but no karyotype × temperature interaction (females: F5,390 = 1.570, P = 0.168; males: F 5,390 = 1.111, P = 0.354).Similar articles
-
The developmental-genetics of canalization.Semin Cell Dev Biol. 2019 Apr;88:67-79. doi: 10.1016/j.semcdb.2018.05.019. Epub 2018 May 24. Semin Cell Dev Biol. 2019. PMID: 29782925 Free PMC article. Review.
-
Symmetry breaking in interspecific Drosophila hybrids is not due to developmental noise.Evolution. 2006 Apr;60(4):746-61. Evolution. 2006. PMID: 16739456
-
Quantitative-genetic analysis of wing form and bilateral asymmetry in isochromosomal lines of Drosophila subobscura using Procrustes methods.J Genet. 2003 Dec;82(3):95-113. doi: 10.1007/BF02715812. J Genet. 2003. PMID: 15133189
-
Control of canalization and evolvability by Hsp90.PLoS One. 2006 Dec 20;1(1):e75. doi: 10.1371/journal.pone.0000075. PLoS One. 2006. PMID: 17183707 Free PMC article.
-
Comparative analysis of morphological traits among Drosophila melanogaster and D. simulans: genetic variability, clines and phenotypic plasticity.Genetica. 2004 Mar;120(1-3):165-79. doi: 10.1023/b:gene.0000017639.62427.8b. Genetica. 2004. PMID: 15088656 Review.
Cited by
-
Development of a Chinese-Indian hybrid (Chindian) rhesus macaque colony at the California National Primate Research Center by introgression.J Med Primatol. 2009 Apr;38(2):86-96. doi: 10.1111/j.1600-0684.2008.00305.x. Epub 2008 Aug 18. J Med Primatol. 2009. PMID: 18715266 Free PMC article.
-
Epidermal growth factor receptor and transforming growth factor-beta signaling contributes to variation for wing shape in Drosophila melanogaster.Genetics. 2006 Jul;173(3):1417-31. doi: 10.1534/genetics.105.053868. Epub 2006 Apr 28. Genetics. 2006. PMID: 16648592 Free PMC article.
-
A single basis for developmental buffering of Drosophila wing shape.PLoS One. 2006 Dec 20;1(1):e7. doi: 10.1371/journal.pone.0000007. PLoS One. 2006. PMID: 17183701 Free PMC article.
-
Multidimensional analysis of Drosophila wing variation in Evolution Canyon.J Genet. 2008 Dec;87(4):407-19. doi: 10.1007/s12041-008-0063-x. J Genet. 2008. PMID: 19147930
-
The developmental-genetics of canalization.Semin Cell Dev Biol. 2019 Apr;88:67-79. doi: 10.1016/j.semcdb.2018.05.019. Epub 2018 May 24. Semin Cell Dev Biol. 2019. PMID: 29782925 Free PMC article. Review.
References
-
- Gibson G, Wagner GP. Canalization in evolutionary theory: a stabilizing theory? Bioessays. 2000;22:372–380. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
