

Surface plasmon resonance (SPR) confirms that MEPE binds to PHEX via the MEPE-ASARM motif: a model for impaired mineralization in X-linked rickets (HYP)
- PMID: 15664000
- PMCID: PMC3361744
- DOI: 10.1016/j.bone.2004.09.015
Surface plasmon resonance (SPR) confirms that MEPE binds to PHEX via the MEPE-ASARM motif: a model for impaired mineralization in X-linked rickets (HYP)
Abstract
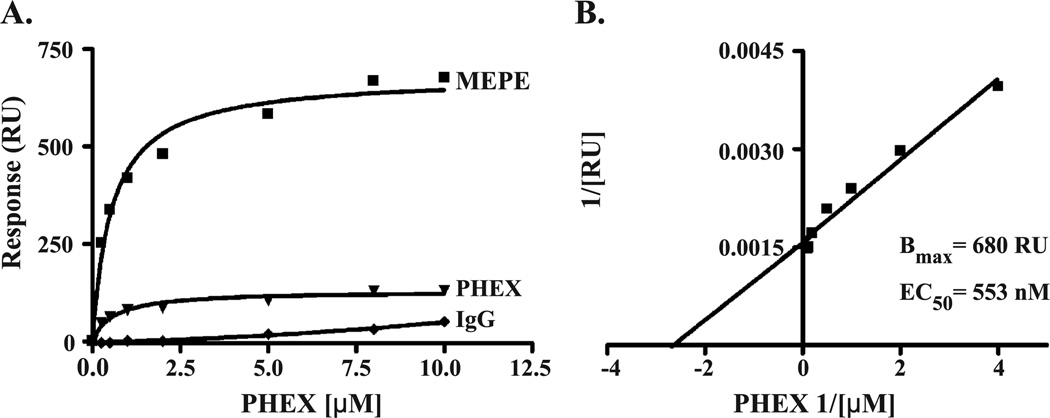
Matrix Extracellular Phospho-glycoprotEin (MEPE) and proteases are elevated and PHEX is defective in HYP. PHEX prevents proteolysis of MEPE and release of a protease-resistant MEPE-ASARM peptide, an inhibitor of mineralization (minhibin). Thus, in HYP, mutated PHEX may contribute to increased ASARM peptide release. Moreover, binding of MEPE by PHEX may regulate this process in normal subjects. The nature of the PHEX-MEPE nonproteolytic interaction(s) (direct or indirect) is/are unknown. Our aims were to determine (1) whether PHEX binds specifically to MEPE, (2) whether the binding involves the ASARM motif region, and (3) whether free ASARM peptide affects mineralization in vivo in mice. Protein interactions between MEPE and recombinant soluble PHEX (secPHEX) were measured using surface plasmon resonance (SPR). Briefly, secPHEX, MEPE, and control protein (IgG) were immobilized on a Biacore CM5 sensor chip, and SPR experiments were performed on a Biacore 3000 high-performance research system. Pure secPHEX was then injected at different concentrations, and interactions with immobilized proteins were measured. To determine MEPE sequences interacting with secPHEX, the inhibitory effects of MEPE-ASARM peptides (phosphorylated and nonphosphorylated), control peptides, and MEPE midregion RGD peptides on secPHEX binding to chip-immobilized MEPE were measured. ASARM peptide and etidronate-mediated mineralization inhibition in vivo and in vitro were determined by quenched calcein fluorescence in hind limbs and calvariae in mice and by histological Sanderson stain. A specific, dose-dependent and Zn-dependent protein interaction between secPHEX and immobilized MEPE occurs (EC50 of 553 nM). Synthetic MEPE PO4-ASARM peptide inhibits the PHEX-MEPE interaction (K(D(app)) = 15 uM and B(max/inhib) = 68%). In contrast, control and MEPE-RGD peptides had no effect. Subcutaneous administration of ASARM peptide resulted in marked quenching of fluorescence in calvariae and hind limbs relative to vehicle controls indicating impaired mineralization. Similar results were obtained with etidronate. Sanderson-stained calvariae also indicated a marked increase in unmineralized osteoid with ASARM peptide and etidronate groups. We conclude that PHEX and MEPE form a nonproteolytic protein interaction via the MEPE carboxy-terminal ASARM motif, and the ASARM peptide inhibits mineralization in vivo. The binding of MEPE and ASARM peptide by PHEX may explain why loss of functional osteoblast-expressed PHEX results in defective mineralization in HYP.
Figures









References
-
- Aisa MC, Beccari T, Costanzi E, Maggio D. Cathepsin B in osteoblasts. Biochim Biophys Acta. 2003;1621:149–159. - PubMed
-
- Argiro L, Desbarats M, Glorieux FH, Ecarot B. Mepe, the gene encoding a tumor-secreted protein in oncogenic hypophosphatemic osteomalacia, is expressed in bone. Genomics. 2001;74:342–351. - PubMed
-
- Bai X, Miao D, Panda D, Grady S, McKee MD, Goltzman D, et al. Partial rescue of the Hyp phenotype by osteoblast-targeted PHEX (phosphate-regulating gene with homologies to endopeptidases on the X chromosome) expression. Mol Endocrinol. 2002;16:2913–2925. - PubMed
-
- Benet-Pages A, Lorenz-Depiereux B, Zischka H, White KE, Econs TM, Strom TM. FGF23 is processed by proprotein convertases but not by PHEX. Bone. 2004;35:455–462. - PubMed
-
- Bianchetti L, Oudet C, Poch O. M13 endopeptidases: new conserved motifs correlated with structure, and simultaneous phylogenetic occurrence of PHEX and the bony fish. Proteins. 2002;47:481–488. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
