

Exposure of acidic residues as a danger signal for recognition of fibrinogen and other macromolecules by integrin alphaXbeta2
- PMID: 15665082
- PMCID: PMC547869
- DOI: 10.1073/pnas.0409057102
Exposure of acidic residues as a danger signal for recognition of fibrinogen and other macromolecules by integrin alphaXbeta2
Abstract
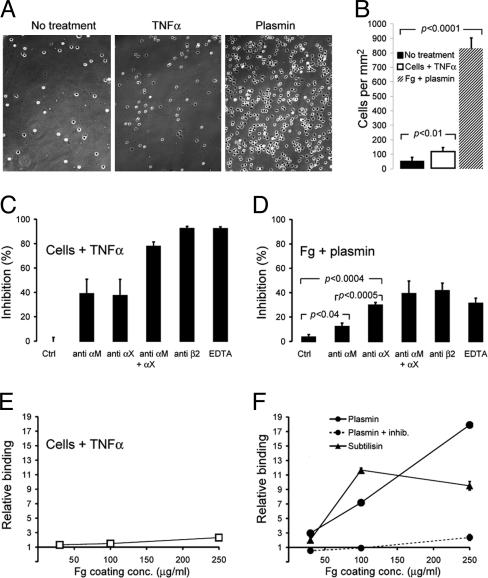
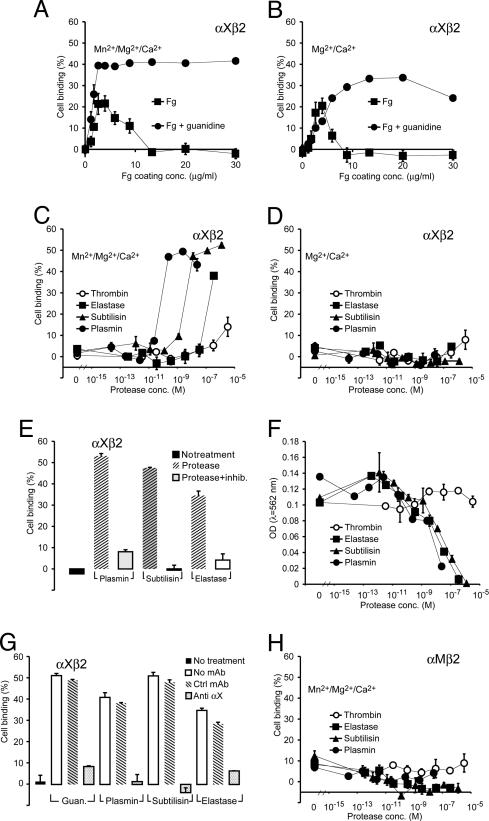
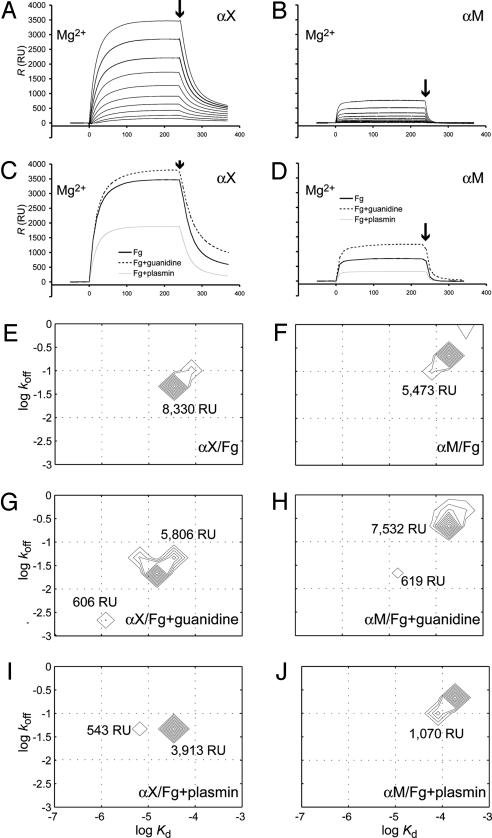
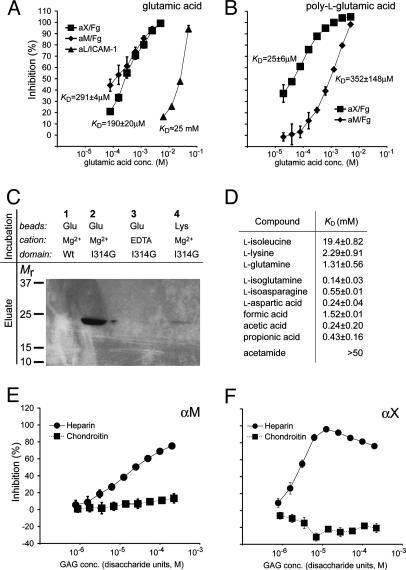
The structural integrity of tissue proteins is damaged in processes ranging from remodeling of the extracellular matrix to destruction by microbial pathogens. Leukocytes play a prominent role in tissue surveillance and repair. However, it remains enigmatic what features of structurally decayed proteins prompt recognition by leukocyte cell-surface receptors. Here, we report that adhesion of human neutrophil granulocytes to fibrinogen is greatly increased by plasmin digestion in a mode where alphaXbeta2 dominates the integrin-dependent binding. The bacterial protease subtilisin also enhances binding by alphaXbeta2. The alphaX ligand binding domain has an unusually high affinity for carboxyl groups, with KD at approximately 100 microM. Our findings implicate enhanced accessibility of negatively charged residues in structurally decayed proteins as a pattern recognition motif for alphaXbeta2 integrin. Comparisons among integrins show relevance of these findings to the large number of ligands recognized by alphaMbeta2 and alphaXbeta2 but not alphaLbeta2. The observations suggest that the pericellular proteolysis at the leading edge of neutrophils not only facilitates passage through the extracellular matrix but also manufactures binding sites for alphaXbeta2.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
