

Transient interference with staphylococcal quorum sensing blocks abscess formation
- PMID: 15665088
- PMCID: PMC547845
- DOI: 10.1073/pnas.0407661102
Transient interference with staphylococcal quorum sensing blocks abscess formation
Abstract
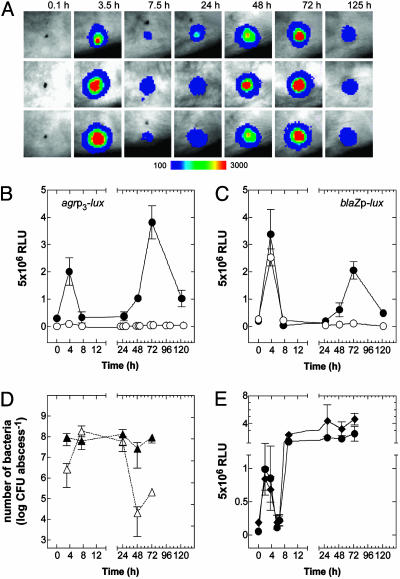
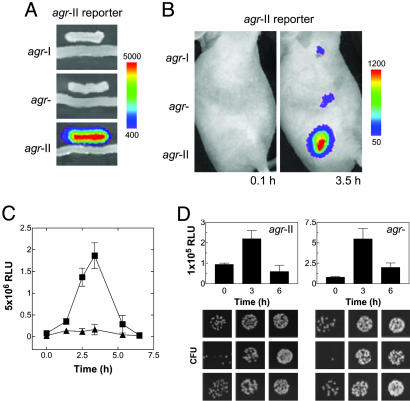
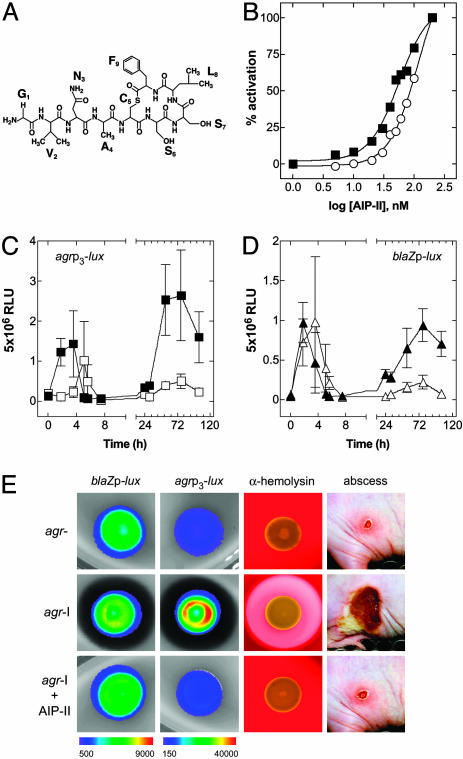
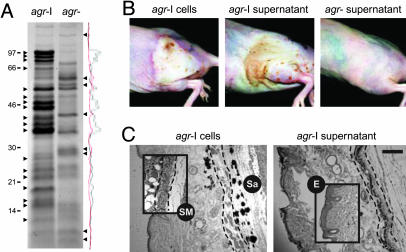
The staphylococcal virulon is controlled largely by the agr locus, a global accessory gene regulator that is autoinduced by a self-coded peptide (AIP) and is therefore a quorum sensor. The agr locus has diverged within and between species, giving rise to AIP variants that inhibit heterologous agr activation, an effect with therapeutic potential against Staphylococcus aureus: a single dose of an inhibitory AIP blocks the formation of an experimental murine abscess. As the AIP is unstable at physiological pH, owing to its essential thiolactone bond, its single-dose efficacy seems paradoxical, which has led us to analyze the in vivo kinetics of agr activation and the consequences of its blockage by a heterologous AIP. Initially, the infecting bacteria grow rapidly, achieving sufficient population density within the first 3 h to activate agr, and then enter a neutrophil-induced metabolic eclipse lasting for 2-3 d, followed by agr reactivation concomitantly with the development of the abscess. The inhibitory AIP prevents agr expression only during its short in vivo lifetime, suggesting that the agr-induced and therefore quorum-dependent synthesis of virulence factors shortly after infection is necessary for the subsequent development of the abscess lesion and bacterial survival. We confirm this finding by showing that a sterile agr+ supernatant causes a sterile abscess similar to the septic abscess caused by live bacteria. These results may provide a biological rationale for regulation of virulence factor expression by quorum sensing rather than by response to specific host signals.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
