

Regulation of choline deficiency apoptosis by epidermal growth factor in CWSV-1 rat hepatocytes
- PMID: 15665516
- PMCID: PMC2424026
- DOI: 10.1159/000083653
Regulation of choline deficiency apoptosis by epidermal growth factor in CWSV-1 rat hepatocytes
Abstract
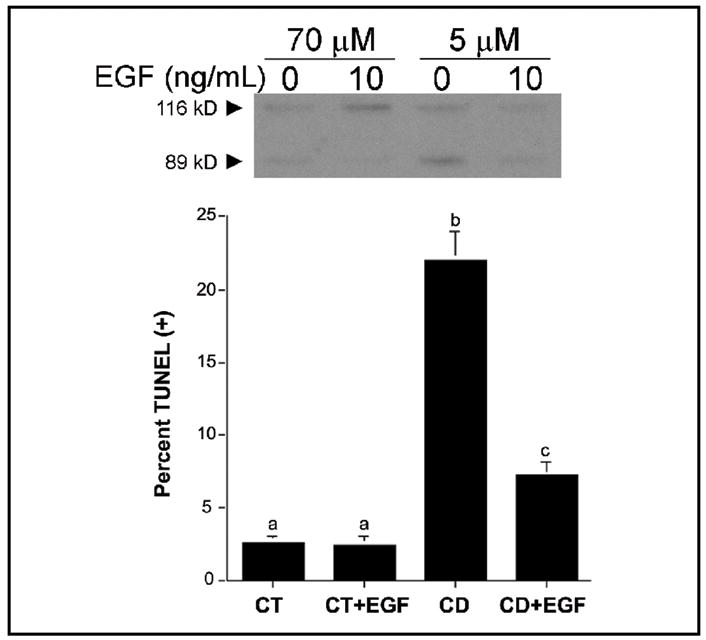
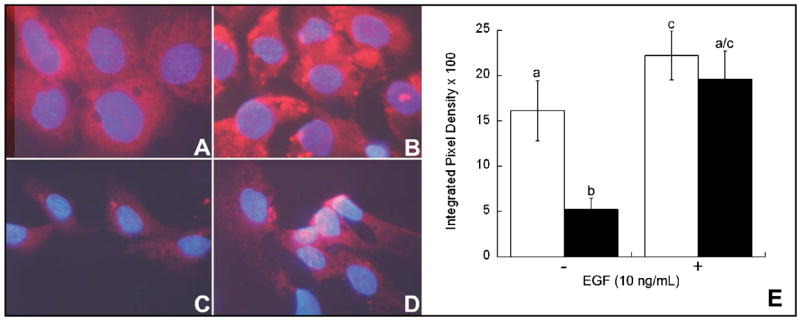
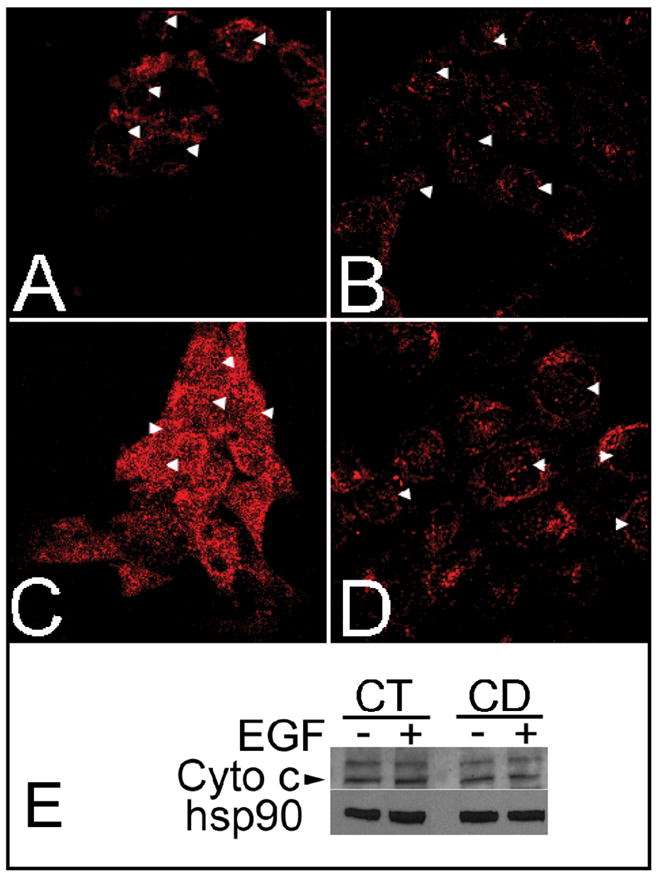
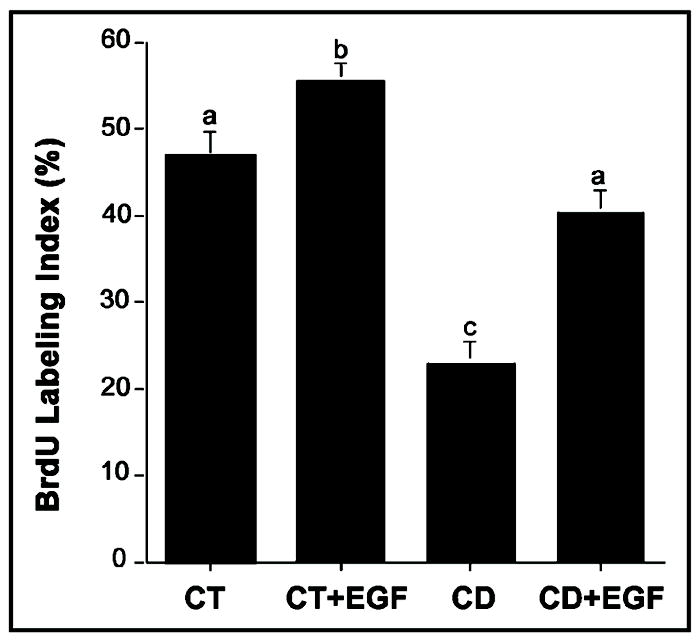
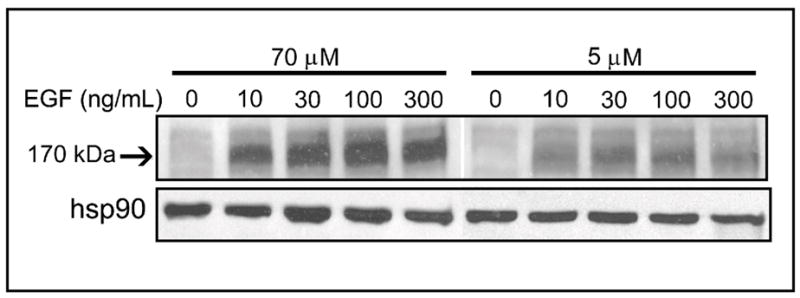
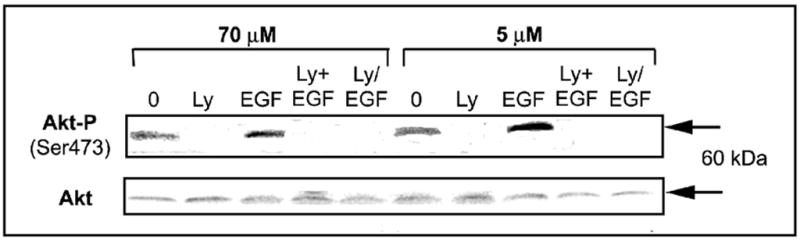
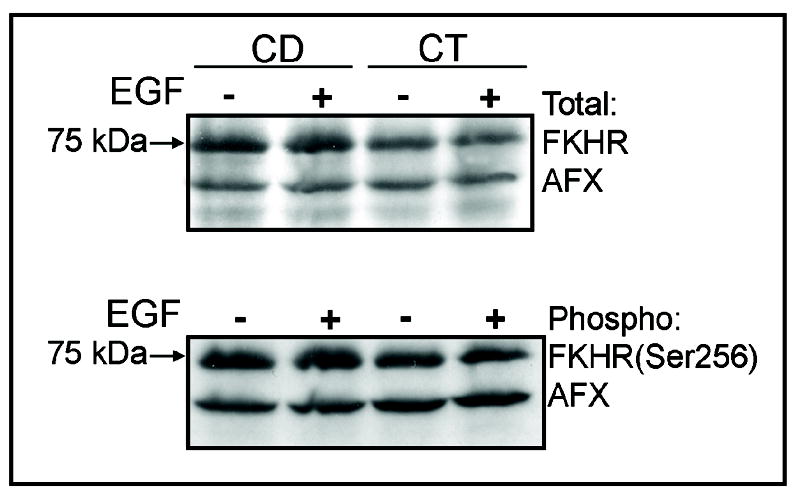
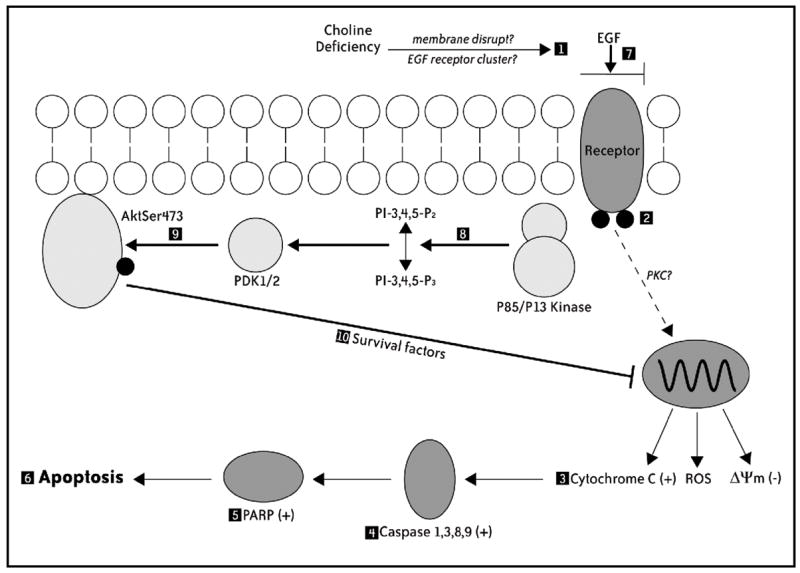
Previous studies show that acute choline deficiency (CD) triggers apoptosis in cultured rat hepatocytes (CWSV-1 cells). We demonstrate that prolonged EGF stimulation (10 ng/mL x 48 hrs) restores cell proliferation, as assessed by BrdU labeling, and protects cells from CD-induced apoptosis, as assessed by TUNEL labeling and cleavage of poly(ADP-ribose) polymerase. However, EGF rescue was not accompanied by restoration of depleted intracellular concentrations of choline, glycerphosphocholine, phosphocholine, or phosphatidylcholine. In contrast, we show that EGF stimulation blocks apoptosis by restoring mitochondrial membrane potential (Delta Psi(m)), as determined using the potential-sensitive dye chloromethyl-X-rosamine, and by preventing the release and nuclear localization of cytochrome c. We investigated whether EGF rescue involves EGF receptor phosphorylation and activation of the down-stream cell survival factor Akt. Compared to cells in control medium (CT, 70 micromol choline x 48 hrs), cells in CD medium (5 micromol choline) were less sensitive to EGF-induced (0-300 ng/mL x 5 min) receptor tyrosine phosphorylation. Compared to cells in CT medium, cells in CD medium treated with EGF (10 ng/mL x 5 min) exhibited higher levels of phosphatidylinositol 3-kinase (PI3K)-dependent phosphorylation of AktSer473. Inactivation of PI3K was sufficient to block EGF-stimulated activation of Akt, restoration of mitochondrial Delta Psi(m), and prevention of cytochrome c release. These studies indicate that stimulation with EGF activates a cell survival response against CD-apoptosis by restoring mitochondrial membrane potential and preventing cytochrome c release and nuclear translocation which are mediated by activation of Akt in hepatocytes.
Copyright 2005 S. Karger AG, Basel.
Figures
References
-
- Food and Nutrition Board, Institute of Medicine. Dietary References Intakes for Thiamin, Riboflavin, Niacin, Vitamin B6, Folate, Vitamin B12, Pantothenic Acid, Biotin and Choline. National Academy Press; Washington, D.C: 1998. - PubMed
-
- Blusztajn JK. Choline: a vital amine. Science. 1998;281:794–795. - PubMed
-
- Jackowski S. Coordination of membrane phospholipid synthesis with the cell cycle. J Biol Chem. 1994;269:3858–3867. - PubMed
-
- Zeisel SH. Choline: essential for brain development and function. Adv Pediatr. 1997;44:263–295. - PubMed
-
- Albright CD, Liu R, Bethea TC, da Costa K-A, Salganik RI, Zeisel SH. Choline deficiency induces apoptosis in CWSV-1 hepatocytes in culture. FASEB J. 1996;10:510–516. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources