

Catalytically inactive human cathepsin D triggers fibroblast invasive growth
- PMID: 15668295
- PMCID: PMC2171724
- DOI: 10.1083/jcb.200403078
Catalytically inactive human cathepsin D triggers fibroblast invasive growth
Abstract
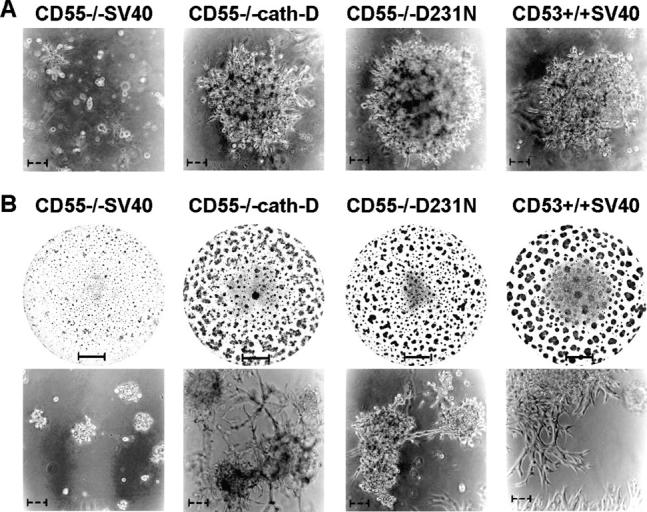
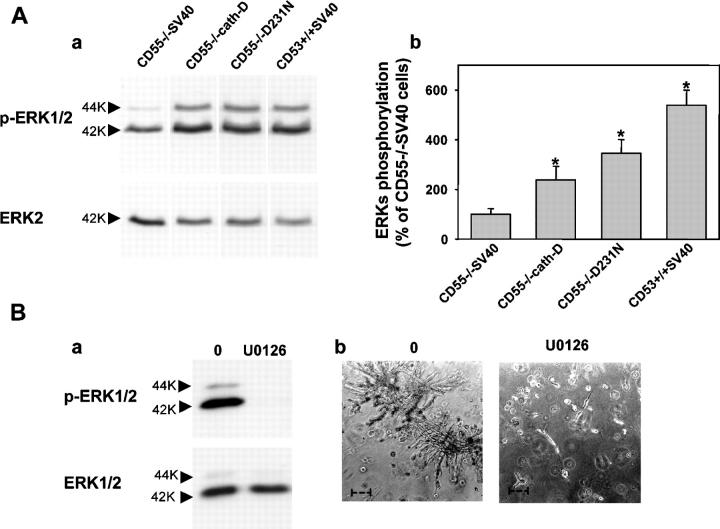
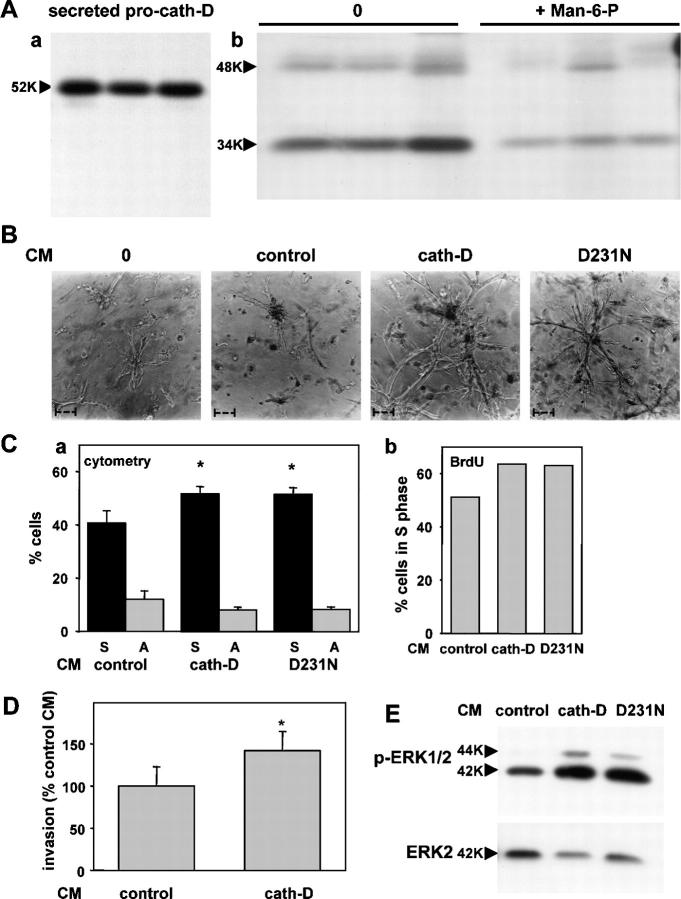
The aspartyl-protease cathepsin D (cath-D) is overexpressed and hypersecreted by epithelial breast cancer cells and stimulates their proliferation. As tumor epithelial-fibroblast cell interactions are important events in cancer progression, we investigated whether cath-D overexpression affects also fibroblast behavior. We demonstrate a requirement of cath-D for fibroblast invasive growth using a three-dimensional (3D) coculture assay with cancer cells secreting or not pro-cath-D. Ectopic expression of cath-D in cath-D-deficient fibroblasts stimulates 3D outgrowth that is associated with a significant increase in fibroblast proliferation, survival, motility, and invasive capacity, accompanied by activation of the ras-MAPK pathway. Interestingly, all these stimulatory effects on fibroblasts are independent of cath-D proteolytic activity. Finally, we show that pro-cath-D secreted by cancer cells is captured by fibroblasts and partially mimics effects of transfected cath-D. We conclude that cath-D is crucial for fibroblast invasive outgrowth and could act as a key paracrine communicator between cancer and stromal cells, independently of its catalytic activity.
Figures



Similar articles
-
The hyaluronan-binding protease upregulates ERK1/2 and PI3K/Akt signalling pathways in fibroblasts and stimulates cell proliferation and migration.Cell Signal. 2005 Dec;17(12):1486-94. doi: 10.1016/j.cellsig.2005.03.007. Epub 2005 Apr 19. Cell Signal. 2005. PMID: 16153533
-
A mutated cathepsin-D devoid of its catalytic activity stimulates the growth of cancer cells.Oncogene. 2001 Oct 18;20(47):6920-9. doi: 10.1038/sj.onc.1204843. Oncogene. 2001. PMID: 11687971
-
Cathepsin D: newly discovered functions of a long-standing aspartic protease in cancer and apoptosis.Cancer Lett. 2006 Jun 18;237(2):167-79. doi: 10.1016/j.canlet.2005.06.007. Epub 2005 Jul 19. Cancer Lett. 2006. PMID: 16046058 Review.
-
Pro-cathepsin D interacts with the extracellular domain of the beta chain of LRP1 and promotes LRP1-dependent fibroblast outgrowth.J Cell Sci. 2010 Oct 1;123(Pt 19):3336-46. doi: 10.1242/jcs.070938. Epub 2010 Sep 7. J Cell Sci. 2010. PMID: 20826454 Free PMC article.
-
Pathophysiological functions of cathepsin D: Targeting its catalytic activity versus its protein binding activity?Biochimie. 2010 Nov;92(11):1635-43. doi: 10.1016/j.biochi.2010.05.009. Epub 2010 May 21. Biochimie. 2010. PMID: 20493920 Review.
Cited by
-
Procathepsin D and cancer: From molecular biology to clinical applications.World J Clin Oncol. 2010 Nov 10;1(1):35-40. doi: 10.5306/wjco.v1.i1.35. World J Clin Oncol. 2010. PMID: 21603309 Free PMC article.
-
SPARC in cancer-associated fibroblasts is an independent poor prognostic factor in non-metastatic triple-negative breast cancer and exhibits pro-tumor activity.Int J Cancer. 2023 Mar 15;152(6):1243-1258. doi: 10.1002/ijc.34345. Epub 2022 Nov 30. Int J Cancer. 2023. PMID: 36346290 Free PMC article.
-
A New Cathepsin D Targeting Drug Delivery System Based on Immunoliposomes Functionalized with Lipidated Pepstatin A.Pharmaceutics. 2023 Oct 14;15(10):2464. doi: 10.3390/pharmaceutics15102464. Pharmaceutics. 2023. PMID: 37896224 Free PMC article.
-
Enzymatically active cathepsin D sensitizes breast carcinoma cells to TRAIL.Tumour Biol. 2016 Aug;37(8):10685-96. doi: 10.1007/s13277-016-4958-5. Epub 2016 Feb 11. Tumour Biol. 2016. PMID: 26867770
-
Yeast as a tool to explore cathepsin D function.Microb Cell. 2015 Jul 11;2(7):225-234. doi: 10.15698/mic2015.07.212. Microb Cell. 2015. PMID: 28357298 Free PMC article. Review.
References
-
- Bar-Sagi, D., and A. Hall. 2000. Ras and Rho GTPases: a family reunion. Cell 103:227–238. - PubMed
-
- Basset, P., J.P. Bellocq, C. Wolf, I. Stoll, P. Hutin, J.M. Limacher, O. Podhajcer, M.P. Chenard, M.C. Rio, and P. Chambon. 1990. A novel metalloproteinase gene specifically expressed in stromal cells of breast carcinomas. Nature. 348:699–704. - PubMed
-
- Berchem, G.J., M. Glondu, M. Gleizes, J.P. Brouillet, F. Vignon, M. Garcia, and E. Liaudet-Coopman. 2002. Cathepsin-D affects multiple steps of tumor progression: Proliferation, angiogenesis and apoptosis. Oncogene. 21:5951–5955. - PubMed
-
- Bidere, N., H.K. Lorenzo, S. Carmona, M. Laforge, F. Harper, C. Dumont, and A. Senik. 2003. Cathepsin D triggers Bax activation, resulting in selective apoptosis-inducing factor (AIF) relocation in T lymphocytes entering the early commitment phase to apoptosis. J. Biol. Chem. 278:31401–31411. - PubMed
-
- Bisson, C., S. Blacher, M. Polette, J.F. Blanc, F. Kebers, J. Desreux, B. Tetu, J. Rosenbaum, J.M. Foidart, P. Birembaut, and A. Noel. 2003. Restricted expression of membrane type 1-matrix metalloproteinase by myofibroblasts adjacent to human breast cancer cells. Int. J. Cancer. 105:7–13. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
