

Genomic instability, endoreduplication, and diminished Ig class-switch recombination in B cells lacking Nbs1
- PMID: 15668392
- PMCID: PMC547831
- DOI: 10.1073/pnas.0406289102
Genomic instability, endoreduplication, and diminished Ig class-switch recombination in B cells lacking Nbs1
Abstract
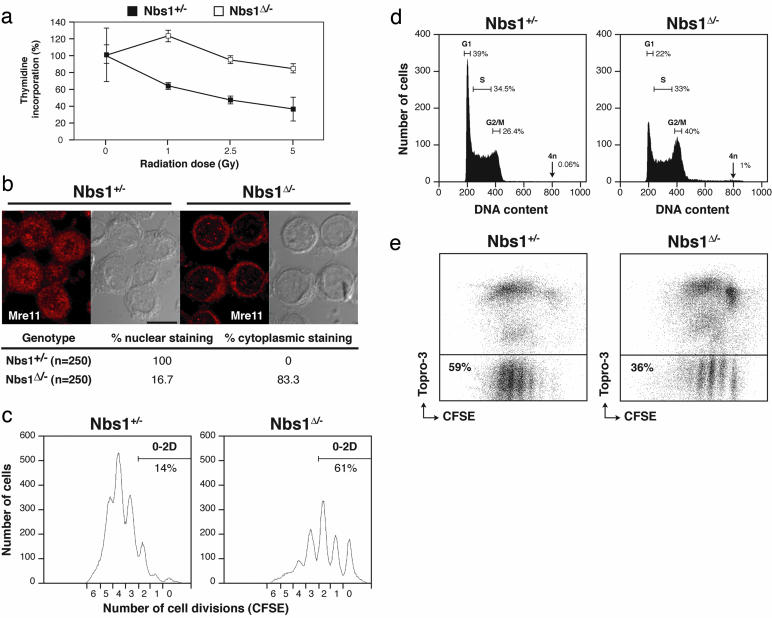
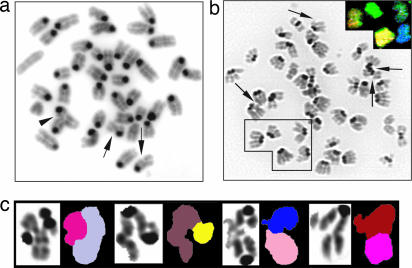
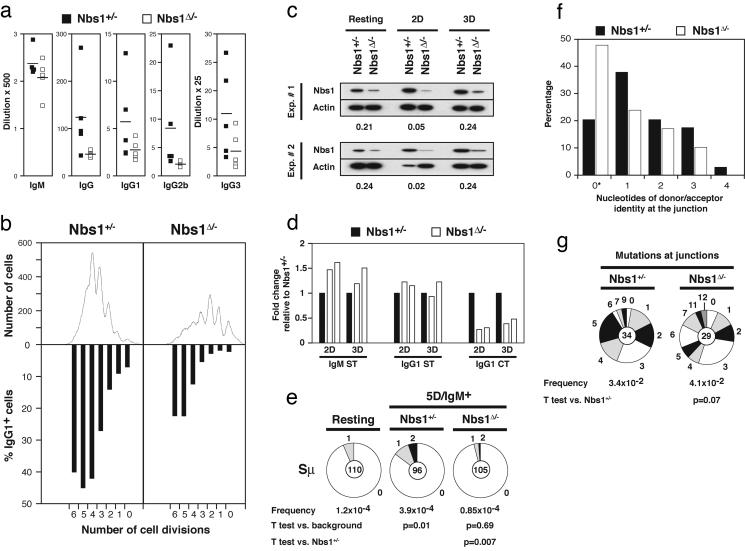
Mre11, Rad50, and Nbs1 form an evolutionarily conserved protein complex (Mre11-Rad50-Nbs1, MRN) that has been proposed to function as a DNA damage sensor. Hypomorphic mutations in Mre11 and Nbs1 result in the human ataxia-telangiectasia-like disorder and Nijmegen breakage syndrome (NBS), respectively. In contrast, complete inactivation of Mre11, Rad50, or Nbs1 leads to early embryonic lethality, suggesting that the hypomorphic mutations may fail to reveal some of the essential functions of MRN. Here, we use Cre-loxP-mediated recombination to restrict Nbs1 deletion to B lymphocytes. We find that disruption of Nbs1 results in the accumulation of high levels of spontaneous DNA damage, impaired proliferation, and chromosomal endoreduplication. Moreover, we show that Ig class-switch recombination (CSR) is diminished in Nbs1-deficient B cells. The CSR defect is B cell-intrinsic, independent of switch-region transcription, and a consequence of inefficient recombination at the DNA level. Our findings reveal that Nbs1 is critical for efficient Ig CSR and maintenance of the integrity of chromosomal structure and number.
Figures
References
-
- Li, Z., Woo, C. J., Iglesias-Ussel, M. D., Ronai, D. & Scharff, M. D. (2004) Genes Dev. 18, 1-11. - PubMed
-
- Chaudhuri, J. & Alt, F. W. (2004) Nat. Rev. Immunol. 4, 541-552. - PubMed
-
- Muramatsu, M., Kinoshita, K., Fagarasan, S., Yamada, S., Shinkai, Y. & Honjo, T. (2000) Cell 102, 553-563. - PubMed
-
- Revy, P., Muto, T., Levy, Y., Geissmann, F., Plebani, A., Sanal, O., Catalan, N., Forveille, M., Dufourcq-Labelouse, R., Gennery, A., et al. (2000) Cell 102, 565-575. - PubMed
-
- Neuberger, M. S., Harris, R. S., Di Noia, J. & Petersen-Mahrt, S. K. (2003) Trends Biochem. Sci. 28, 305-312. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
