

Polymodal sensory function of the Caenorhabditis elegans OCR-2 channel arises from distinct intrinsic determinants within the protein and is selectively conserved in mammalian TRPV proteins
- PMID: 15673683
- PMCID: PMC6725639
- DOI: 10.1523/JNEUROSCI.3107-04.2005
Polymodal sensory function of the Caenorhabditis elegans OCR-2 channel arises from distinct intrinsic determinants within the protein and is selectively conserved in mammalian TRPV proteins
Abstract
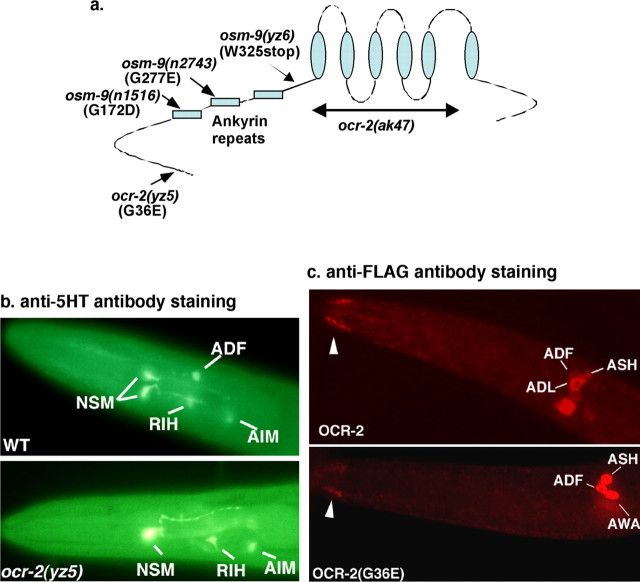
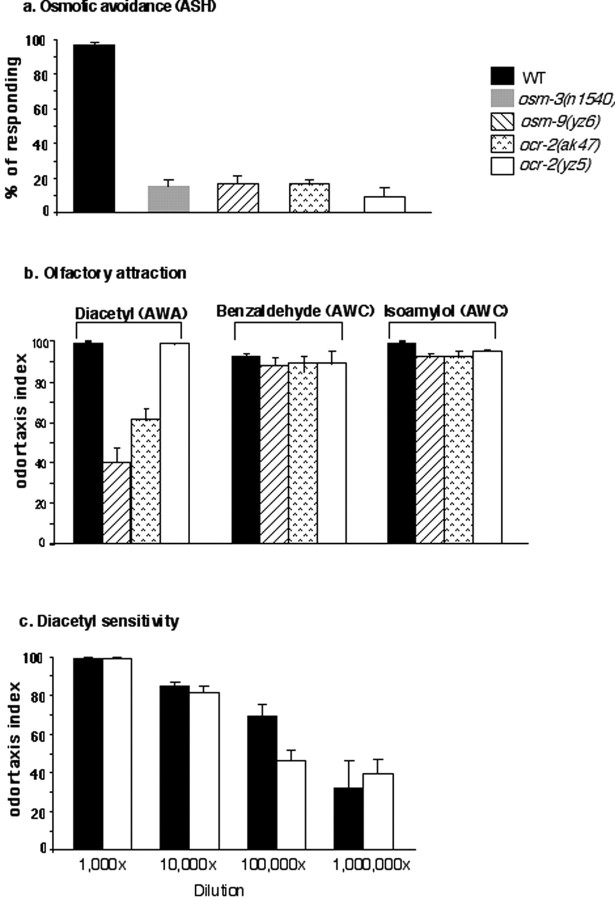
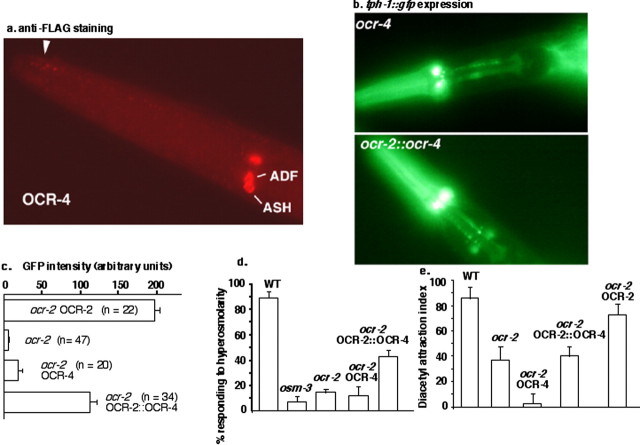
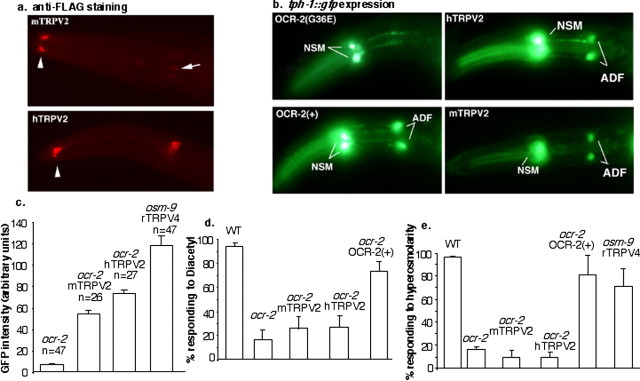
Caenorhabditis elegans OCR-2 (OSM-9 and capsaicin receptor-related) is a TRPV (vanilloid subfamily of transient receptor potential channel) protein that regulates serotonin (5-HT) biosynthesis in chemosensory neurons and also mediates olfactory and osmotic sensation. Here, we identify the molecular basis for the polymodal function of OCR-2 in its native cellular environment. We show that OCR-2 function in 5-HT production and osmotic sensing is governed by its N-terminal region upstream of the ankyrin repeats domain, but the diacetyl sensitivity is mediated by independent mechanisms. The ocr-2(yz5) mutation results in a glycine-to-glutamate substitution (G36E) within the N-terminal region. The G36E substitution causes dramatic downregulation of 5-HT synthesis in the ADF neurons, eliminates osmosensation mediated by the ASH neurons, but does not affect the response to the odorant diacetyl mediated by the AWA neurons. Conversely, wild-type sequence of the N-terminal segment confers osmotic sensitivity and upregulation of 5-HT production to a normally insensitive C. elegans homolog, OCR-4, but this chimeric channel does not respond to diacetyl stimuli. Furthermore, expression of either the mouse or human TRPV2 gene under the ocr-2 promoter can substantially restore 5-HT biosynthesis in ocr-2-null mutants but cannot improve the deficits in osmotic or olfactory sensation, suggesting that TRPV2 can substitute for the role of OCR-2 only in serotonergic neurons. Thus, different sensory functions of OCR-2 arise from separable intrinsic determinants, and specific functional properties of TRPV channel proteins may be selectively conserved across phyla.
Figures

Similar articles
-
Caenorhabditis elegans TRPV ion channel regulates 5HT biosynthesis in chemosensory neurons.Development. 2004 Apr;131(7):1629-38. doi: 10.1242/dev.01047. Epub 2004 Mar 3. Development. 2004. PMID: 14998926
-
OSM-9, a novel protein with structural similarity to channels, is required for olfaction, mechanosensation, and olfactory adaptation in Caenorhabditis elegans.J Neurosci. 1997 Nov 1;17(21):8259-69. doi: 10.1523/JNEUROSCI.17-21-08259.1997. J Neurosci. 1997. PMID: 9334401 Free PMC article.
-
Mammalian TRPV4 (VR-OAC) directs behavioral responses to osmotic and mechanical stimuli in Caenorhabditis elegans.Proc Natl Acad Sci U S A. 2003 Nov 25;100 Suppl 2(Suppl 2):14531-6. doi: 10.1073/pnas.2235619100. Epub 2003 Oct 27. Proc Natl Acad Sci U S A. 2003. PMID: 14581619 Free PMC article.
-
The mechanosensitive nature of TRPV channels.Pflugers Arch. 2005 Oct;451(1):193-203. doi: 10.1007/s00424-005-1424-4. Epub 2005 May 21. Pflugers Arch. 2005. PMID: 15909178 Review.
-
Function and regulation of TRP family channels in C. elegans.Pflugers Arch. 2009 Sep;458(5):851-60. doi: 10.1007/s00424-009-0678-7. Epub 2009 May 8. Pflugers Arch. 2009. PMID: 19421772 Free PMC article. Review.
Cited by
-
Cilia and cell cycle re-entry: more than a coincidence.Cell Cycle. 2011 Aug 15;10(16):2683-90. doi: 10.4161/cc.10.16.17009. Epub 2011 Aug 15. Cell Cycle. 2011. PMID: 21814045 Free PMC article. Review.
-
What do we know about the transient receptor potential vanilloid 2 (TRPV2) ion channel?FEBS J. 2013 Nov;280(21):5471-87. doi: 10.1111/febs.12302. Epub 2013 May 28. FEBS J. 2013. PMID: 23615321 Free PMC article. Review.
-
Dependence of regulatory volume decrease on transient receptor potential vanilloid 4 (TRPV4) expression in human corneal epithelial cells.Cell Calcium. 2008 Oct;44(4):374-85. doi: 10.1016/j.ceca.2008.01.008. Cell Calcium. 2008. PMID: 18355916 Free PMC article.
-
TRPV4 plays an evolutionary conserved role in the transduction of osmotic and mechanical stimuli in live animals.J Physiol. 2005 Aug 15;567(Pt 1):53-8. doi: 10.1113/jphysiol.2005.088963. Epub 2005 Jun 16. J Physiol. 2005. PMID: 15961428 Free PMC article. Review.
-
Molecular and cellular limits to somatosensory specificity.Mol Pain. 2008 Apr 18;4:14. doi: 10.1186/1744-8069-4-14. Mol Pain. 2008. PMID: 18419827 Free PMC article. Review.
References
-
- Allessandri-Haber N, Yeh JJ, Boyd AE, Parada CA, Chen X, Reichling DB, Levine JD (2003) Hypotonicity induces TRPV4-mediated nociception in rat. Neuron 39: 497-511. - PubMed
-
- Baldi P, Pollastri G (2003) The principled design of large-scale recursive neural network architectures-DAG-RNNs and the protein structure prediction problem. J Machine Learning Res 4: 575-602.
-
- Baldi P, Brunak S, Frasconi P, Soda G, Pollastri G (1999) Exploiting the past and the future in protein secondary structure prediction. Bioinformatics 15: 937-946. - PubMed
-
- Bargmann CI, Horvitz HR (1991a) Chemosensory neurons with overlapping functions direct chemotaxis to multiple chemicals in C. elegans Neuron 5: 729-742. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous