

Functional interaction of reverse gyrase with single-strand binding protein of the archaeon Sulfolobus
- PMID: 15673717
- PMCID: PMC548347
- DOI: 10.1093/nar/gki202
Functional interaction of reverse gyrase with single-strand binding protein of the archaeon Sulfolobus
Abstract
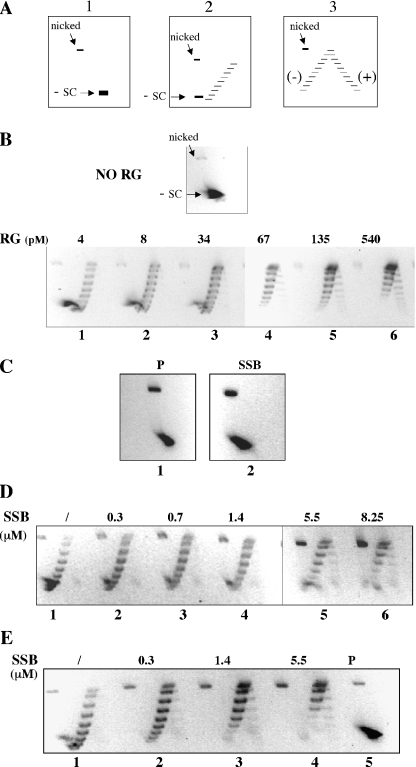
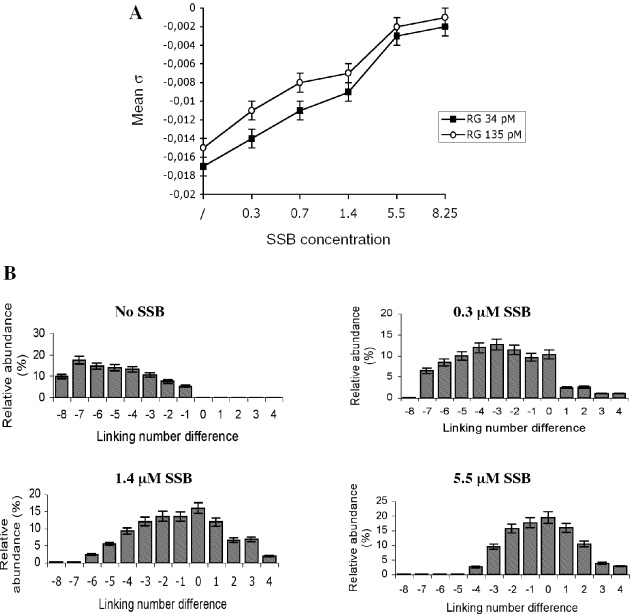
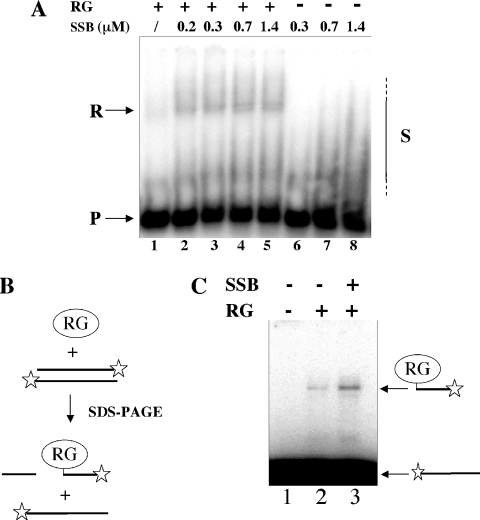
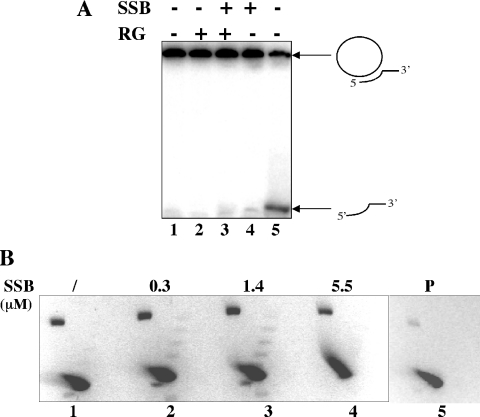
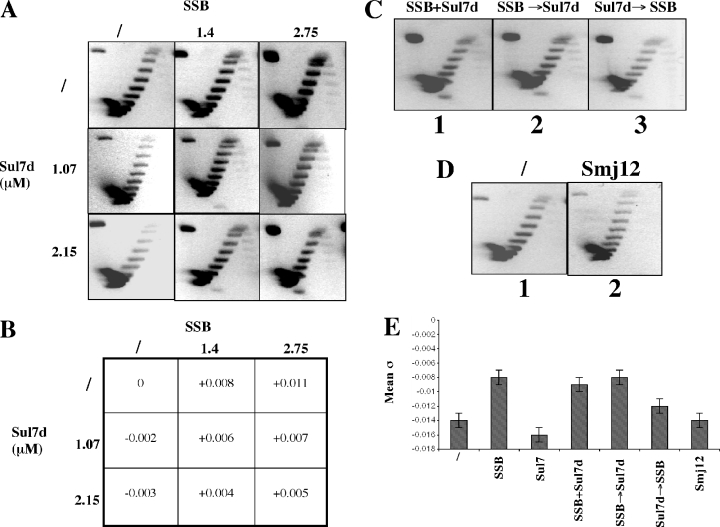
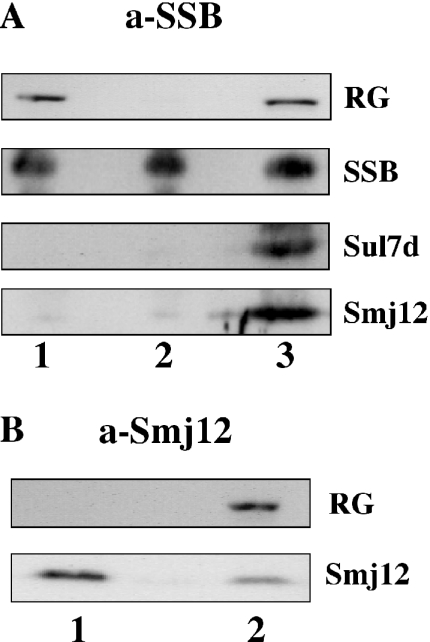
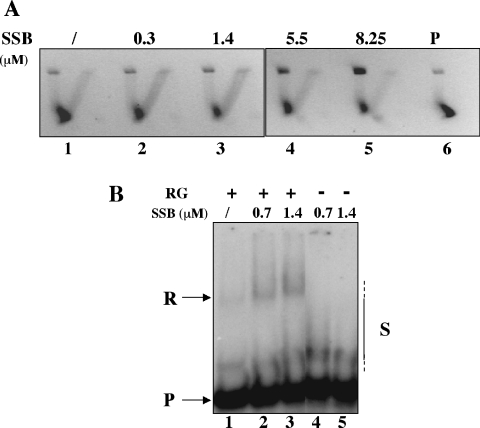
Reverse gyrase is a unique hyperthermophile-specific DNA topoisomerase that induces positive supercoiling. It is a modular enzyme composed of a topoisomerase IA and a helicase domain, which cooperate in the ATP-dependent positive supercoiling reaction. Although its physiological function has not been determined, it can be hypothesized that, like the topoisomerase-helicase complexes found in every organism, reverse gyrase might participate in different DNA transactions mediated by multiprotein complexes. Here, we show that reverse gyrase activity is stimulated by the single-strand binding protein (SSB) from the archaeon Sulfolobus solfataricus. Using a combination of in vitro assays we analysed each step of the complex reverse gyrase reaction. SSB stimulates all the steps of the reaction: binding to DNA, DNA cleavage, strand passage and ligation. By co-immunoprecipitation of cell extracts we show that reverse gyrase and SSB assemble a complex in the presence of DNA, but do not make stable protein-protein interactions. In addition, SSB stimulates reverse gyrase positive supercoiling activity on DNA templates associated with the chromatin protein Sul7d. Furthermore, SSB enhances binding and cleavage of UV-irradiated substrates by reverse gyrase. The results shown here suggest that these functional interactions may have biological relevance and that the interplay of different DNA binding proteins might modulate reverse gyrase activity in DNA metabolic pathways.
Figures
Similar articles
-
Adenosine 5'-O-(3-thio)triphosphate (ATPgammaS) promotes positive supercoiling of DNA by T. maritima reverse gyrase.J Mol Biol. 2007 Aug 3;371(1):197-209. doi: 10.1016/j.jmb.2007.05.031. Epub 2007 May 18. J Mol Biol. 2007. PMID: 17560602
-
Reverse gyrase: an unusual DNA manipulator of hyperthermophilic organisms.Ital J Biochem. 2007 Jun;56(2):103-9. Ital J Biochem. 2007. PMID: 17722650 Review.
-
Physical and functional interaction between archaeal single-stranded DNA-binding protein and the 5'-3' nuclease NurA.Biochem Biophys Res Commun. 2008 Mar 14;367(3):523-9. doi: 10.1016/j.bbrc.2007.10.019. Epub 2007 Oct 15. Biochem Biophys Res Commun. 2008. PMID: 18194801
-
Influence of chromatin and single strand binding proteins on the activity of an archaeal MCM.J Mol Biol. 2006 Apr 14;357(5):1345-50. doi: 10.1016/j.jmb.2006.01.074. Epub 2006 Feb 6. J Mol Biol. 2006. PMID: 16490210
-
Reverse gyrase and genome stability in hyperthermophilic organisms.Biochem Soc Trans. 2009 Feb;37(Pt 1):69-73. doi: 10.1042/BST0370069. Biochem Soc Trans. 2009. PMID: 19143604 Review.
Cited by
-
Binding dynamics of a monomeric SSB protein to DNA: a single-molecule multi-process approach.Nucleic Acids Res. 2015 Dec 15;43(22):10907-24. doi: 10.1093/nar/gkv1225. Epub 2015 Nov 17. Nucleic Acids Res. 2015. PMID: 26578575 Free PMC article.
-
Archaea: A Gold Mine for Topoisomerase Diversity.Front Microbiol. 2021 May 25;12:661411. doi: 10.3389/fmicb.2021.661411. eCollection 2021. Front Microbiol. 2021. PMID: 34113328 Free PMC article. Review.
-
Structure and Properties of DNA Molecules Over The Full Range of Biologically Relevant Supercoiling States.Sci Rep. 2018 Apr 18;8(1):6163. doi: 10.1038/s41598-018-24499-5. Sci Rep. 2018. PMID: 29670174 Free PMC article.
-
Insight into the cellular involvement of the two reverse gyrases from the hyperthermophilic archaeon Sulfolobus solfataricus.BMC Mol Biol. 2014 Sep 9;15:18. doi: 10.1186/1471-2199-15-18. BMC Mol Biol. 2014. PMID: 25200003 Free PMC article.
-
Genome stability: recent insights in the topoisomerase reverse gyrase and thermophilic DNA alkyltransferase.Extremophiles. 2014 Sep;18(5):895-904. doi: 10.1007/s00792-014-0662-9. Epub 2014 Aug 8. Extremophiles. 2014. PMID: 25102812 Review.
References
-
- Champoux J.J. DNA topoisomerases: structure, function, and mechanism. Annu. Rev. Biochem. 2001;70:369–413. - PubMed
-
- Wang J.C. Cellular roles of DNA topoisomerases: a molecular perspective. Nature Rev. Mol. Cell Biol. 2002;3:430–440. - PubMed
-
- Duguet M. Reverse gyrase. In: Eckstein F., Lilley D.M.J., editors. Nucleic Acids and Molecular Biology. Vol. IX. Berlin, Germany: Springer-Verlag; 1995. pp. 84–114.
-
- Forterre P., Bergerat A., Lopez-Garcia P. The unique DNA topology and DNA topoisomerases of hyperthermophilic archaea. FEMS Microbiol. Rev. 1996;18:237–248. - PubMed
-
- Forterre P. A hot story from comparative genomics: reverse gyrase is the only hyperthermophile-specific protein. Trends Genet. 2002;18:236–237. - PubMed
