

Assignment of an essential role for the Neurospora frequency gene in circadian entrainment to temperature cycles
- PMID: 15677317
- PMCID: PMC548525
- DOI: 10.1073/pnas.0406506102
Assignment of an essential role for the Neurospora frequency gene in circadian entrainment to temperature cycles
Abstract
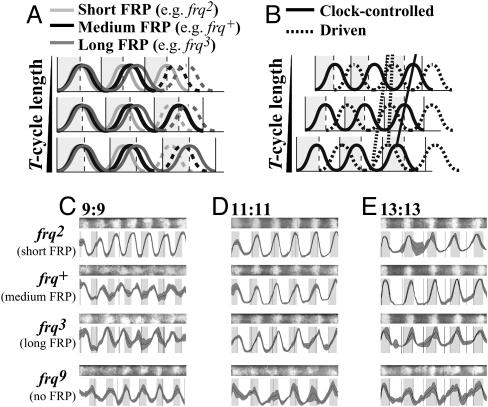
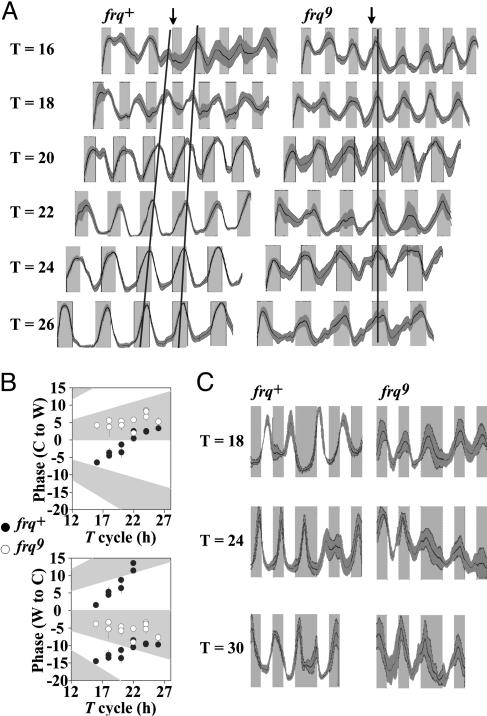
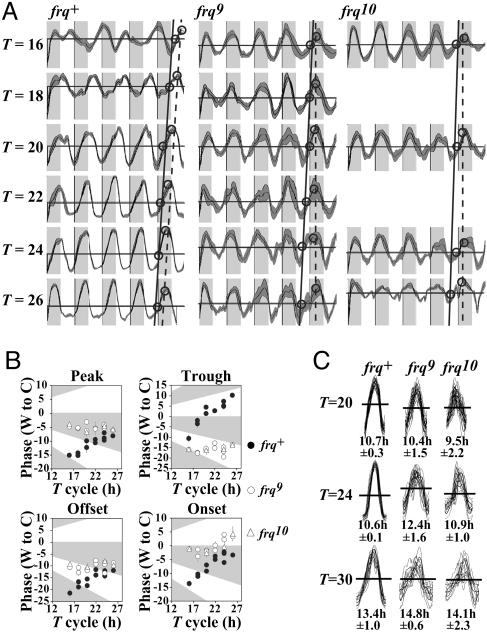
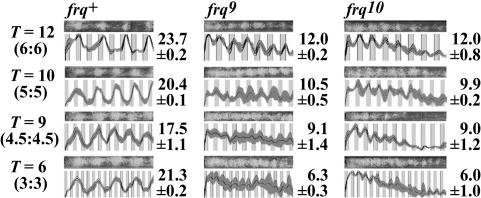
Circadian systems include slave oscillators and central pacemakers, and the cores of eukaryotic circadian clocks described to date are composed of transcription and translation feedback loops (TTFLs). In the model system Neurospora, normal circadian rhythmicity requires a TTFL in which a White Collar complex (WCC) activates expression of the frequency (frq) gene, and the FRQ protein feeds back to attenuate that activation. To further test the centrality of this TTFL to the circadian mechanism in Neurospora, we used low-amplitude temperature cycles to compare WT and frq-null strains under conditions in which a banding rhythm was elicited. WT cultures were entrained to these temperature cycles. Unlike those normal strains, however, frq-null mutants did not truly entrain to the same cycles. Their peaks and troughs always occurred in the cold and warm periods, respectively, strongly suggesting that the rhythm in Neurospora lacking frq function simply is driven by the temperature cycles. Previous reports suggested that a FRQ-less oscillator (FLO) could be entrained to temperature cycles, rather than being driven, and speculated that the FLO was the underlying circadian-rhythm generator. These inferences appear to derive from the use of a phase reference point affected by both the changing waveform and the phase of the oscillation. Examination of several other phase markers as well as results of additional experimental tests indicate that the FLO is, at best, a slave oscillator to the TTFL, which underlies circadian rhythm generation in Neurospora.
Figures
Similar articles
-
Circadian clock genes frequency and white collar-1 are not essential for entrainment to temperature cycles in Neurospora crassa.Proc Natl Acad Sci U S A. 2006 Mar 21;103(12):4469-74. doi: 10.1073/pnas.0510404103. Epub 2006 Mar 14. Proc Natl Acad Sci U S A. 2006. PMID: 16537415 Free PMC article.
-
PRD-1, a Component of the Circadian System of Neurospora crassa, Is a Member of the DEAD-box RNA Helicase Family.J Biol Rhythms. 2016 Jun;31(3):258-71. doi: 10.1177/0748730416639717. Epub 2016 Mar 29. J Biol Rhythms. 2016. PMID: 27029286
-
Effects of prd circadian clock mutations on FRQ-less rhythms in Neurospora.J Biol Rhythms. 2010 Apr;25(2):71-80. doi: 10.1177/0748730409360889. J Biol Rhythms. 2010. PMID: 20348458
-
The molecular workings of the Neurospora biological clock.Novartis Found Symp. 2003;253:184-98; discussion 102-9, 198-202, 281-4. Novartis Found Symp. 2003. PMID: 14712922 Review.
-
The neurospora circadian system.J Biol Rhythms. 2004 Oct;19(5):414-24. doi: 10.1177/0748730404269116. J Biol Rhythms. 2004. PMID: 15534321 Review.
Cited by
-
CCA1 alternative splicing as a way of linking the circadian clock to temperature response in Arabidopsis.Plant Signal Behav. 2012 Sep 1;7(9):1194-6. doi: 10.4161/psb.21300. Epub 2012 Aug 17. Plant Signal Behav. 2012. PMID: 22899064 Free PMC article.
-
Circadian rhythms from multiple oscillators: lessons from diverse organisms.Nat Rev Genet. 2005 Jul;6(7):544-56. doi: 10.1038/nrg1633. Nat Rev Genet. 2005. PMID: 15951747 Free PMC article. Review.
-
Functional analysis of 110 phosphorylation sites on the circadian clock protein FRQ identifies clusters determining period length and temperature compensation.G3 (Bethesda). 2023 Feb 9;13(2):jkac334. doi: 10.1093/g3journal/jkac334. G3 (Bethesda). 2023. PMID: 36537198 Free PMC article.
-
Circadian output, input, and intracellular oscillators: insights into the circadian systems of single cells.Cold Spring Harb Symp Quant Biol. 2007;72:201-14. doi: 10.1101/sqb.2007.72.067. Cold Spring Harb Symp Quant Biol. 2007. PMID: 18419278 Free PMC article. Review.
-
The COP9 signalosome regulates the Neurospora circadian clock by controlling the stability of the SCFFWD-1 complex.Genes Dev. 2005 Jul 1;19(13):1518-31. doi: 10.1101/gad.1322205. Epub 2005 Jun 16. Genes Dev. 2005. PMID: 15961524 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
