

Homeostatic capabilities of the choroid plexus epithelium in Alzheimer's disease
- PMID: 15679944
- PMCID: PMC546405
- DOI: 10.1186/1743-8454-1-3
Homeostatic capabilities of the choroid plexus epithelium in Alzheimer's disease
Abstract
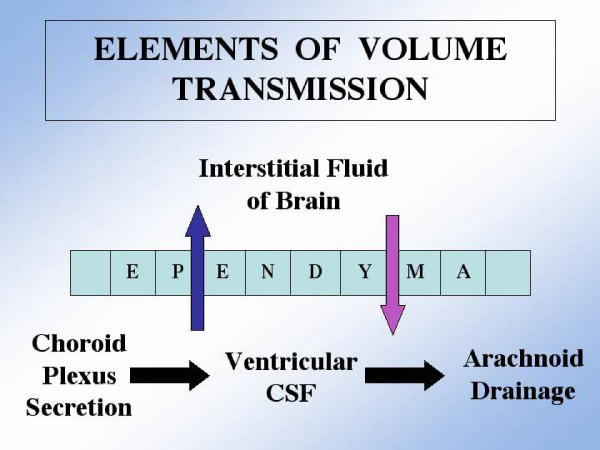
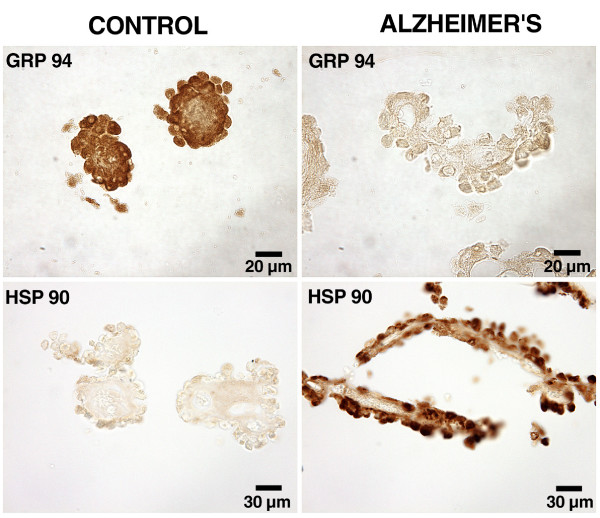
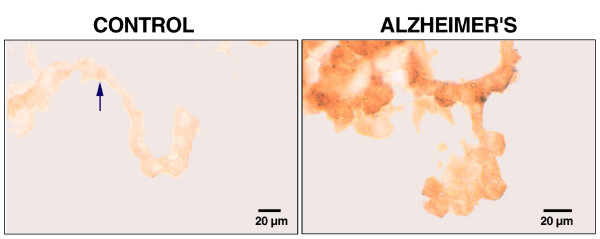
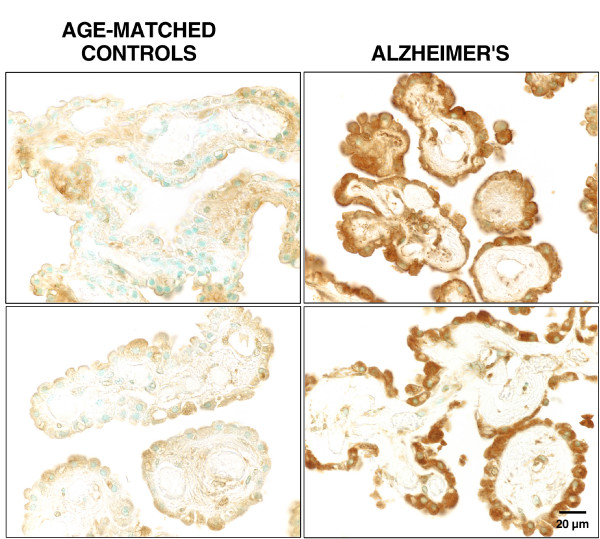
As the secretory source of vitamins, peptides and hormones for neurons, the choroid plexus (CP) epithelium critically provides substances for brain homeostasis. This distributive process of cerebrospinal fluid (CSF) volume transmission reaches many cellular targets in the CNS. In ageing and ageing-related dementias, the CP-CSF system is less able to regulate brain interstitial fluid. CP primarily generates CSF bulk flow, and so its malfunctioning exacerbates Alzheimers disease (AD). Considerable attention has been devoted to the blood-brain barrier in AD, but more insight is needed on regulatory systems at the human blood-CSF barrier in order to improve epithelial function in severe disease. Using autopsied CP specimens from AD patients, we immunocytochemically examined expression of heat shock proteins (HSP90 and GRP94), fibroblast growth factor receptors (FGFr) and a fluid-regulatory protein (NaK2Cl cotransporter isoform 1 or NKCC1). CP upregulated HSP90, FGFr and NKCC1, even in end-stage AD. These CP adjustments involve growth factors and neuropeptides that help to buffer perturbations in CNS water balance and metabolism. They shed light on CP-CSF system responses to ventriculomegaly and the altered intracranial pressure that occurs in AD and normal pressure hydrocephalus. The ability of injured CP to express key regulatory proteins even at Braak stage V/VI, points to plasticity and function that may be boosted by drug treatment to expedite CSF dynamics. The enhanced expression of human CP 'homeostatic proteins' in AD dementia is discussed in relation to brain deficits and pharmacology.
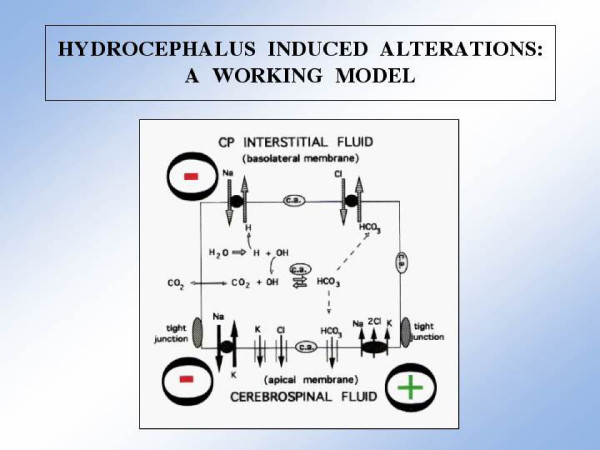
Figures
Similar articles
-
Comparative transcriptomics of choroid plexus in Alzheimer's disease, frontotemporal dementia and Huntington's disease: implications for CSF homeostasis.Fluids Barriers CNS. 2018 May 31;15(1):18. doi: 10.1186/s12987-018-0102-9. Fluids Barriers CNS. 2018. PMID: 29848382 Free PMC article.
-
Choroid plexus genes for CSF production and brain homeostasis are altered in Alzheimer's disease.Fluids Barriers CNS. 2018 Dec 12;15(1):34. doi: 10.1186/s12987-018-0120-7. Fluids Barriers CNS. 2018. PMID: 30541599 Free PMC article.
-
The choroid plexus-cerebrospinal fluid system: from development to aging.Curr Top Dev Biol. 2005;71:1-52. doi: 10.1016/S0070-2153(05)71001-2. Curr Top Dev Biol. 2005. PMID: 16344101 Review.
-
Enhanced prospects for drug delivery and brain targeting by the choroid plexus-CSF route.Pharm Res. 2005 Jul;22(7):1011-37. doi: 10.1007/s11095-005-6039-0. Epub 2005 Jul 22. Pharm Res. 2005. PMID: 16028003 Review.
-
Traumatic brain injury and recovery mechanisms: peptide modulation of periventricular neurogenic regions by the choroid plexus-CSF nexus.J Neural Transm (Vienna). 2011 Jan;118(1):115-33. doi: 10.1007/s00702-010-0498-0. Epub 2010 Oct 10. J Neural Transm (Vienna). 2011. PMID: 20936524 Free PMC article. Review.
Cited by
-
The regulation of brain states by neuroactive substances distributed via the cerebrospinal fluid; a review.Cerebrospinal Fluid Res. 2010 Jan 6;7:1. doi: 10.1186/1743-8454-7-1. Cerebrospinal Fluid Res. 2010. PMID: 20157443 Free PMC article.
-
Antimicrobial peptide β-defensin-1 expression is upregulated in Alzheimer's brain.J Neuroinflammation. 2013 Oct 18;10:127. doi: 10.1186/1742-2094-10-127. J Neuroinflammation. 2013. PMID: 24139179 Free PMC article.
-
Brain uptake of nonsteroidal anti-inflammatory drugs: ibuprofen, flurbiprofen, and indomethacin.Pharm Res. 2006 May;23(5):873-81. doi: 10.1007/s11095-006-9905-5. Epub 2006 May 2. Pharm Res. 2006. PMID: 16715377
-
Cerebral nitric oxide represses choroid plexus NFκB-dependent gateway activity for leukocyte trafficking.EMBO J. 2015 Jul 2;34(13):1816-28. doi: 10.15252/embj.201591468. Epub 2015 May 4. EMBO J. 2015. PMID: 25940071 Free PMC article.
-
Spatial and temporal morphological changes in the subarachnoid space after graded spinal cord contusion in the rat.J Neurotrauma. 2013 Jun 15;30(12):1084-91. doi: 10.1089/neu.2012.2764. J Neurotrauma. 2013. PMID: 23472674 Free PMC article.
References
-
- Johanson CE, Silverberg GD, Donahue JE, Duncan JA, Stopa EG. Choroid plexus and CSF in Alzheimer's Disease: Altered expression and transport of proteins and peptides. In: Zheng W, Chodobski A, editor. The Blood-Cerebrospinal Fluid Barrier. Boca Raton: CRC Press LLC; 2005. pp. 307–339.
-
- Huber JD, Egleton RD, Davis TP. Molecular physiology and pathophysiology of tight junctions in the blood-brain barrier. Trends Neurosci. 2001;24:719–25. - PubMed
-
- Rubenstein E. Relationship of senescence of cerebrospinal fluid circulatory system to dementias of the aged. Lancet. 1998;351:283–285. - PubMed
-
- Silverberg GD, Heit G, Huhn S, Jaffe RA, Chang SD, Bronte-Stewart H, Rubenstein E, Possin K, Saul TA. The cerebrospinal fluid production rate is reduced in dementia of the Alzheimer's type. Neurology. 2001;57:1763–1766. - PubMed
-
- Johanson C. The choroid plexus-CSF nexus: gateway to the brain. In: Conn PM, editor. Neuroscience in Medicine. Humana Press; 2003. pp. 165–195.
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
