

Engineered allosteric ribozymes that respond to specific divalent metal ions
- PMID: 15681614
- PMCID: PMC548338
- DOI: 10.1093/nar/gki182
Engineered allosteric ribozymes that respond to specific divalent metal ions
Abstract
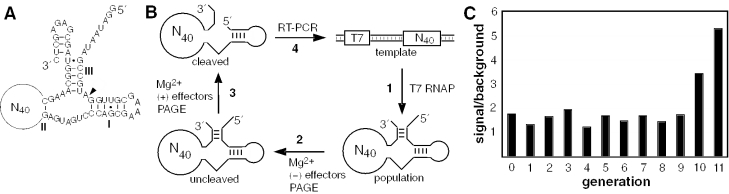
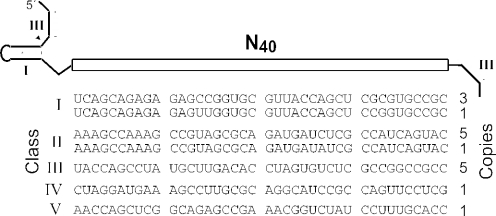
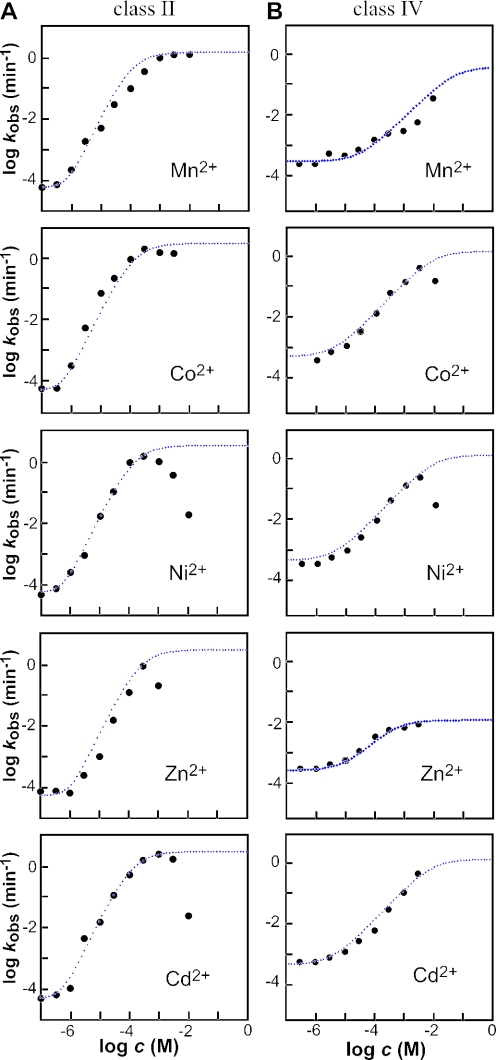
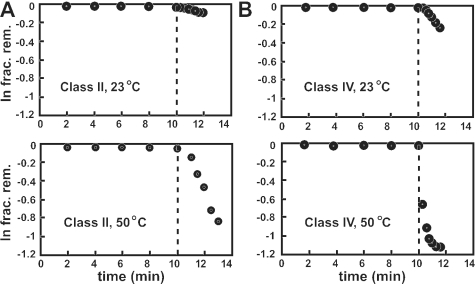
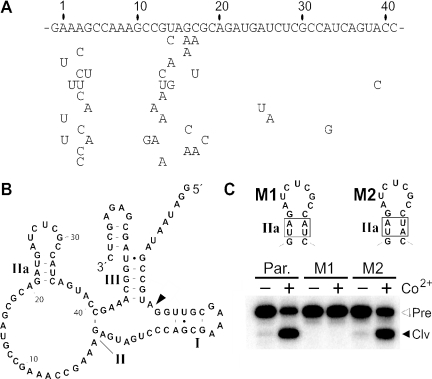
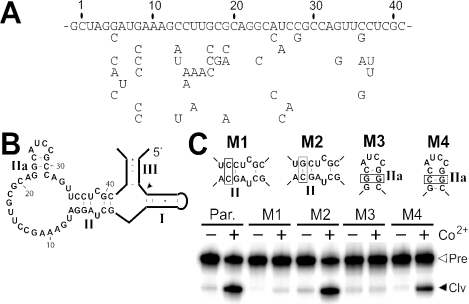
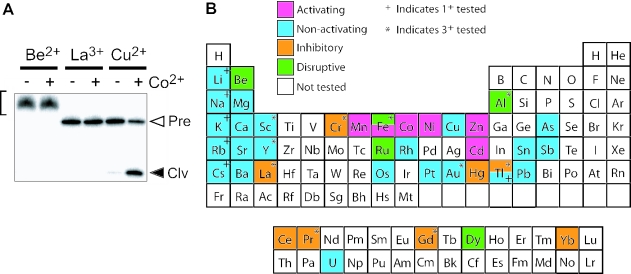
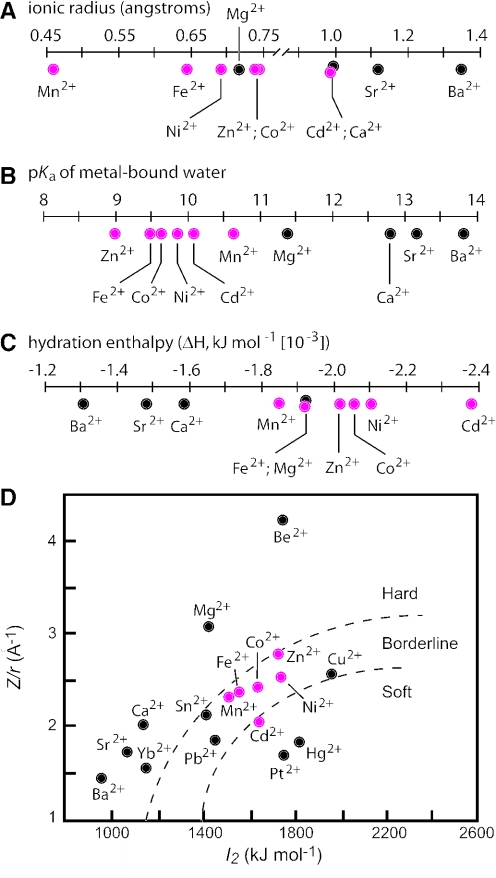
In vitro selection was used to isolate five classes of allosteric hammerhead ribozymes that are triggered by binding to certain divalent metal ion effectors. Each of these ribozyme classes are similarly activated by Mn2+, Fe2+, Co2+, Ni2+, Zn2+ and Cd2+, but their allosteric binding sites reject other divalent metals such as Mg2+, Ca2+ and Sr2+. Through a more comprehensive survey of cations, it was determined that some metal ions (Be2+, Fe3+, Al3+, Ru2+ and Dy2+) are extraordinarily disruptive to the RNA structure and function. Two classes of RNAs examined in greater detail make use of conserved nucleotides within the large internal bulges to form critical structures for allosteric function. One of these classes exhibits a metal-dependent increase in rate constant that indicates a requirement for the binding of two cation effectors. Additional findings suggest that, although complex allosteric functions can be exhibited by small RNAs, larger RNA molecules will probably be required to form binding pockets that are uniquely selective for individual cation effectors.
Figures
Similar articles
-
Effects of divalent metal ions on individual steps of the Tetrahymena ribozyme reaction.Biochemistry. 1997 Jul 8;36(27):8293-303. doi: 10.1021/bi9700678. Biochemistry. 1997. PMID: 9204875
-
Allosteric selection of ribozymes that respond to the second messengers cGMP and cAMP.Nat Struct Biol. 1999 Nov;6(11):1062-71. doi: 10.1038/14947. Nat Struct Biol. 1999. PMID: 10542100
-
Examination of the catalytic fitness of the hammerhead ribozyme by in vitro selection.RNA. 1997 Aug;3(8):914-25. RNA. 1997. PMID: 9257650 Free PMC article.
-
Engineering allosteric regulation into biological catalysts.Chembiochem. 2009 Dec 14;10(18):2824-35. doi: 10.1002/cbic.200900590. Chembiochem. 2009. PMID: 19937897 Review.
-
RNA as a catalyst: natural and designed ribozymes.Bioessays. 1993 May;15(5):299-307. doi: 10.1002/bies.950150503. Bioessays. 1993. PMID: 8343141 Review.
Cited by
-
Identification of the Same Na(+)-Specific DNAzyme Motif from Two In Vitro Selections Under Different Conditions.J Mol Evol. 2015 Dec;81(5-6):225-34. doi: 10.1007/s00239-015-9715-7. Epub 2015 Nov 17. J Mol Evol. 2015. PMID: 26577294
-
Fast production of homogeneous recombinant RNA--towards large-scale production of RNA.Nucleic Acids Res. 2012 Jul;40(13):e102. doi: 10.1093/nar/gks292. Epub 2012 Mar 28. Nucleic Acids Res. 2012. PMID: 22457065 Free PMC article.
-
Simple fluorescent sensors engineered with catalytic DNA 'MgZ' based on a non-classic allosteric design.PLoS One. 2007 Nov 21;2(11):e1224. doi: 10.1371/journal.pone.0001224. PLoS One. 2007. PMID: 18030352 Free PMC article.
-
The importance of peripheral sequences in determining the metal selectivity of an in vitro-selected Co(2+) -dependent DNAzyme.Chembiochem. 2012 Feb 13;13(3):381-91. doi: 10.1002/cbic.201100724. Epub 2012 Jan 17. Chembiochem. 2012. PMID: 22250000 Free PMC article.
-
Activity, folding and Z-DNA formation of the 8-17 DNAzyme in the presence of monovalent ions.J Am Chem Soc. 2009 Apr 22;131(15):5506-15. doi: 10.1021/ja8082939. J Am Chem Soc. 2009. PMID: 19326878 Free PMC article.
References
-
- Patel D.J., Suri A.K. Structure, recognition and discrimination in RNA aptamer complexes with cofactors, amino acids, drugs and aminoglycoside antibiotics. J. Biotechnol. 2000;74:39–60. - PubMed
-
- Brody E.N., Gold L. Aptamers as therapeutic and diagnostic agents. J. Biotechnol. 2000;74:5–13. - PubMed
-
- Koizumi M., Soukup G.A., Kerr J.N., Breaker R.R. Allosteric selection of ribozymes that respond to the second messengers cGMP and cAMP. Nature Struct. Biol. 1999;6:1062–1071. - PubMed
-
- Sassanfar M., Szostak J.W. An RNA motif that binds ATP. Nature. 1993;364:550–553. - PubMed
-
- Lorsch J.R., Szostak J.W. In vitro selection of RNA aptamers specific for cyanocobalamin. Biochemistry. 1994;33:973–982. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
