

The fragile X mental retardation protein and group I metabotropic glutamate receptors regulate levels of mRNA granules in brain
- PMID: 15684045
- PMCID: PMC548595
- DOI: 10.1073/pnas.0409803102
The fragile X mental retardation protein and group I metabotropic glutamate receptors regulate levels of mRNA granules in brain
Abstract
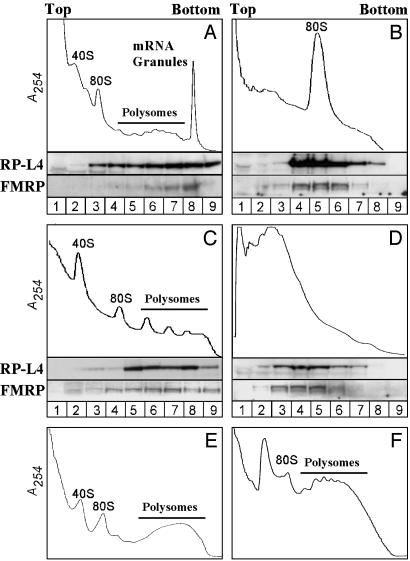
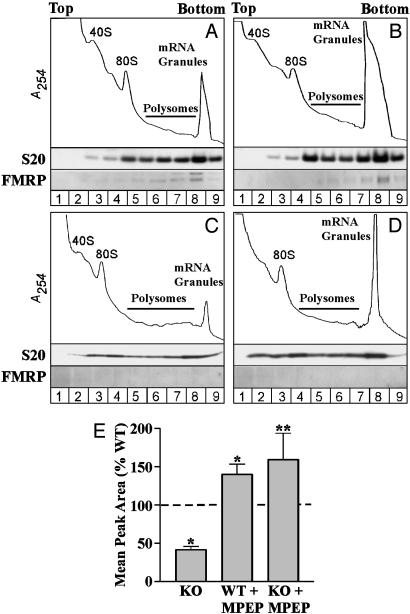
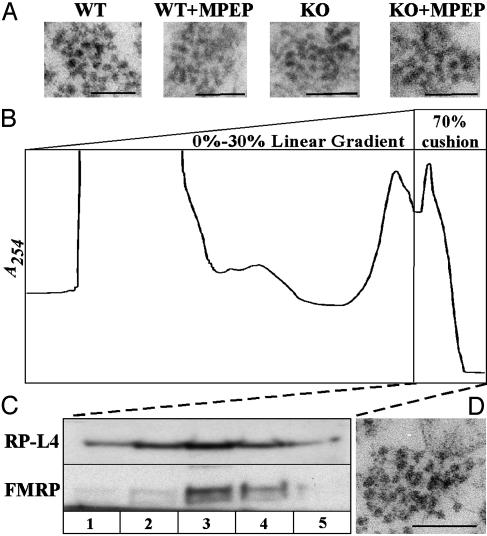
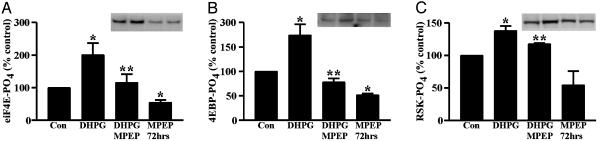
Fragile X syndrome results from the transcriptional silencing of a gene, Fmr1, that codes for an mRNA-binding protein (fragile X mental retardation protein, FMRP) present in neuronal dendrites. FMRP can act as a translational suppressor, and its own translation in dendrites is regulated by group I metabotropic glutamate receptors (mGluRs). Multiple lines of evidence suggest that mGluR-induced translation is exaggerated in Fragile X syndrome because of a lack of translational inhibition normally provided by FMRP. We characterized the role of FMRP in the regulation of mRNA granules, which sediment as a heavy peak after polysomes on sucrose gradients. In WT mouse brain, FMRP distributed with polysomes and granules. EM and biochemical analyses suggested that the granule fraction itself contained clusters of polysomes. In Fmr1 knockout brain, we observed a significant decrease in the amount of mRNA granules relative to WT mice. This difference appeared to be due to a role of FMRP in regulating the activation of granules during mGluR-induced translation; in vivo administration of the mGluR5 antagonist 2-methyl-6-(phenylethynyl)pyridine increased granule content in Fmr1 knockout mouse brain to levels comparable with those seen in WT brain. In accord with a role of mGluR5 in the regulation of ongoing translation in vivo, we observed that the phosphorylation of several initiation factors in response to application of the mGluR1/5 agonist S-3,5-dihydroxyphenylglycine in vitro was blocked by methyl-6-(phenylethynyl)pyridine. Together, these data suggest that although large, polysome-containing granules can form in the absence of FMRP, their use in response to mGluR-induced translation is exaggerated.
Figures
References
-
- O'Donnell, W. T. & Warren, S. T. (2002) Annu. Rev. Neurosci. 25, 315-338. - PubMed
-
- Hagerman, R. J., Amiri, K. & Cronister, A. (1991) Am. J. Med. Genet. 38, 283-287. - PubMed
-
- Hinton, V. J., Brown, W. T., Wisniewski, K. & Rudelli, R. D. (1991) Am. J. Med. Genet. 41, 289-294. - PubMed
-
- Irwin, S. A., Galvez, R. & Greenough, W. T. (2000) Cereb. Cortex 10, 1038-1044. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
