

Involvement of human MCM8 in prereplication complex assembly by recruiting hcdc6 to chromatin
- PMID: 15684404
- PMCID: PMC548026
- DOI: 10.1128/MCB.25.4.1560-1568.2005
Involvement of human MCM8 in prereplication complex assembly by recruiting hcdc6 to chromatin
Abstract
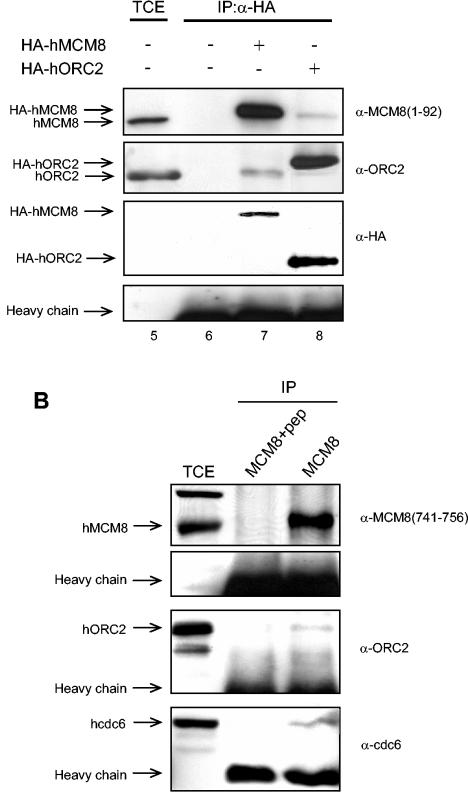
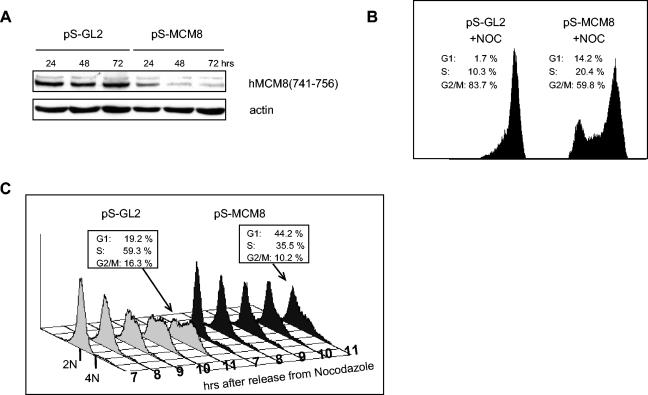
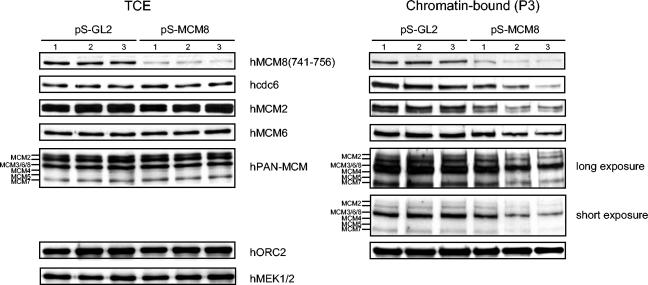
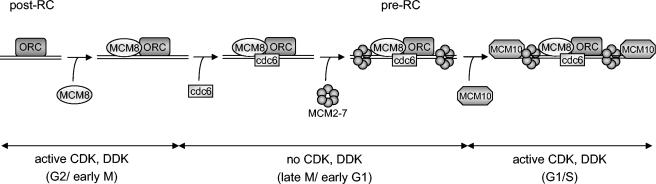
The MCM2-MCM7 complex is an essential component of the prereplication complex (pre-RC), which is recruited by the cdc6 and cdt1 proteins to origins of DNA replication during G(1) phase. Here, we report that the accumulation on chromatin of another member of the MCM protein family, human MCM8 (hMCM8), occurs during early G(1) phase, before the hMCM2-hMCM7 complex binds. hMCM8 interacts in vivo with two components of the pre-RC, namely, hcdc6 and hORC2. Depletion of endogenous hMCM8 protein by RNA interference leads to a delay of entry into S phase, suggesting a role for hMCM8 in G(1) progression. Furthermore, down-regulation of hMCM8 also leads to a reduced loading of hcdc6 and the hMCM2-hMCM7 complex on chromatin. These results suggest that hMCM8 is a crucial component of the pre-RC and that the interaction between hMCM8 and hcdc6 is required for pre-RC assembly.
Figures
Similar articles
-
MCM9 binds Cdt1 and is required for the assembly of prereplication complexes.Mol Cell. 2008 Jul 25;31(2):190-200. doi: 10.1016/j.molcel.2008.07.001. Mol Cell. 2008. PMID: 18657502
-
Colocalization of MCM8 and MCM7 with proteins involved in distinct aspects of DNA replication.Microsc Res Tech. 2008 Apr;71(4):288-97. doi: 10.1002/jemt.20553. Microsc Res Tech. 2008. PMID: 18072282
-
Identification and functional characterization of a new member of the human Mcm protein family: hMcm8.Nucleic Acids Res. 2003 Jan 15;31(2):570-9. doi: 10.1093/nar/gkg136. Nucleic Acids Res. 2003. PMID: 12527764 Free PMC article.
-
Cdt1 and geminin in DNA replication initiation.Subcell Biochem. 2012;62:71-87. doi: 10.1007/978-94-007-4572-8_5. Subcell Biochem. 2012. PMID: 22918581 Review.
-
[Emerging roles for components of pre-replication complexes in chromosomal dynamics].Tanpakushitsu Kakusan Koso. 2009 Mar;54(4 Suppl):531-6. Tanpakushitsu Kakusan Koso. 2009. PMID: 21089504 Review. Japanese. No abstract available.
Cited by
-
Genome wide comparative comprehensive analysis of Plasmodium falciparum MCM family with human host.Commun Integr Biol. 2012 Nov 1;5(6):607-15. doi: 10.4161/cib.21759. Commun Integr Biol. 2012. PMID: 23336032 Free PMC article.
-
Differential requirements for MCM proteins in DNA replication in Drosophila S2 cells.PLoS One. 2007 Sep 5;2(9):e833. doi: 10.1371/journal.pone.0000833. PLoS One. 2007. PMID: 17786205 Free PMC article.
-
Novel biallelic MCMDC2 variants were associated with meiotic arrest and nonobstructive azoospermia.Asian J Androl. 2025 Mar 1;27(2):268-275. doi: 10.4103/aja202495. Epub 2025 Jan 10. Asian J Androl. 2025. PMID: 39789727 Free PMC article.
-
HROB Is Implicated in DNA Replication.Genes (Basel). 2024 Dec 10;15(12):1587. doi: 10.3390/genes15121587. Genes (Basel). 2024. PMID: 39766854 Free PMC article.
-
MCM8 promotes gastric cancer progression through RPS15A and predicts poor prognosis.Cancer Med. 2024 Jul;13(13):e7424. doi: 10.1002/cam4.7424. Cancer Med. 2024. PMID: 38988047 Free PMC article.
References
-
- Aparicio, O. M., D. M. Weinstein, and S. P. Bell. 1997. Components and dynamics of DNA replication complexes in S. cerevisiae: redistribution of MCM proteins and Cdc45p during S phase. Cell 91:59-69. - PubMed
-
- Aves, S. J., N. Tongue, A. J. Foster, and E. A. Hart. 1998. The essential Schizosaccharomyces pombe cdc23 DNA replication gene shares structural and functional homology with the Saccharomyces cerevisiae DNA43 (MCM10) gene. Curr. Genet. 34:164-171. - PubMed
-
- Bell, S. P., and A. Dutta. 2002. DNA replication in eukaryotic cells. Annu. Rev. Biochem. 71:333-374. - PubMed
-
- Bell, S. P., and B. Stillman. 1992. ATP-dependent recognition of eukaryotic origins of DNA replication by a multiprotein complex. Nature 357:128-134. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous